Биогенетический закон — это… Что такое Биогенетический закон?
Биогенетический закон Геккеля-Мюллера (также известен под названиями «закон Геккеля», «закон Мюллера-Геккеля», «закон Дарвина-Мюллера-Геккеля», «основной биогенетический закон»): каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом (филогенез)[1].
Зародыши по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию ГеккеляСыграл важную роль в истории развития науки, однако в настоящее время в своем исходном виде не признается современной биологической наукой.[2][3] По современной трактовке биогенетического закона, предложенной русским биологом А. Н. Северцовым в начале 20 века, в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей.[4]
История создания
Фактически «биогенетический закон» был сформулирован ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции
В 1824—1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своем эмбриональном развитии повторяет взрослые формы более примитивных животных[источник не указан 469 дней].
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своем развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла (он до конца жизни так и не принял эволюционного учения Дарвина), однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» (см. Макроэволюция) и свидетельство происхождения животных одного типа от общего предка.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован (довольно нечётко) английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиностном его состоянии, всех членов одного и того же большого класса» (Дарвин Ч. Соч. М.-Л., 1939, т. 3, с. 636.)
За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер[6]. В своей книге «За Дарвина» (Für Darwin), изданной в 1864 году, он выделяет курсивом мысль: «историческое развитие вида будет отражаться в истории его индивидуального развития».
Краткая афористичная формулировка этого закона была дана немецким естествоиспытателем Эрнстом Геккелем в 1866 г. Краткая формулировка закона звучит следующим образом:
Примеры выполнения биогенетического закона
Яркий пример выполнения биогенетического закона — развитие лягушки, включающее в себя стадию головастика, который по своему строению гораздо больше похож на рыб, чем на земноводных:
У головастика, как и у низших рыб и рыбьих мальков, основой скелета служит хорда, только впоследствии в туловищной части обрастающая хрящевыми позвонками. Череп у головастика хрящевой, и к нему примыкают хорошо развитые хрящевые дуги; дыхание жаберное. Кровеносная система также построена по рыбьему типу: предсердие ещё не разделилось на правую и левую половины, кровь в сердце поступает только венозная, а оттуда через артериальный ствол идёт к жабрам. Если бы развитие головастика остановилось на этой стадии и не шло дальше, мы должны были бы без всяких колебаний отнести такое животное к надклассу рыб.
[7]
Зародыши не только земноводных, но и всех без исключения позвоночных животных также имеют на ранних стадиях развития жаберные щели, двухкамерное сердце и другие признаки, характерные для рыб. Например, птичий зародыш в первые дни насиживания также представляет собой хвостатое рыбообразное существо с жаберными щелями. На этой стадии будущий птенец обнаруживает сходство и с низшими рыбами, и с личинками амфибий, и с ранними стадиями развития других позвоночных животных (в том числе и человека
И пока у куриного зародыша до конца первой недели и задние, и передние конечности имеют вид одинаковых лапок, пока хвост ещё не успел исчезнуть, а из сосочков ещё не сформировались перья, он по всем своим признакам стоит ближе к пресмыкающимся, чем к взрослым птицам.[7]
Зародыш человека в ходе эмбриогенеза проходит через аналогичные стадии. Затем, за период примерно между четвертой и шестой неделями развития он превращается из рыбоподобного организма в организм, неотличимый от зародыша обезьяны, и только потом приобретает человеческие черты.
Такое повторение признаков предков в ходе индивидуального развития особи Геккель назвал рекапитуляция.
Существует множество других примеров рекапитуляций, которые подтверждают выполнение «биогенетического закона» в некоторых случаях. Так, при размножении наземного рака-отшельника пальмового вора его самки перед вылуплением личинок заходят в море, и там из яиц выходят планктонные креветкообразные личинки зоэа, имеющие вполне симметричное брюшко. Затем они превращаются в глаукотоэ и оседают на дно, где находят подходящие раковины брюхоногих моллюсков. Некоторое время они ведут образ жизни, характерный для большинства раков-отшельников, и на этой стадии имеют характерное для этой группы мягкое спиральное брюшко с асимметричными конечностями и дышат жабрами. Выросшие до определенных размеров пальмовые воры покидают раковину, выходят на сушу, приобретают жесткое укороченное брюшко, похожее на абдомен крабов, и навсегда утрачивают способность дышать в воде.
Столь полное выполнение биогенетического закона возможно в тех случаях, когда эволюция онтогенеза происходит путем его удлинения — «надставки стадий»:
- a1 — a2
- a1′ — a2′ — a3′
- a1″ — a2″ — a3″ — a4″
(На этой схеме сверху вниз расположены виды-предки и виды-потомки, а слева направо — стадии их онтогенеза.)
Факты, противоречащие биогенетическому закону
Уже в XIX веке было известно достаточно фактов, противоречащих биогенетическому закону. Так, были известны многочисленные примеры неотении, при которой в ходе эволюции происходит укорочение онтогенеза и выпадение его конечных стадий. В случае неотении взрослая стадия вида-потомка напоминает личиночную стадию вида-предка, а не наоборот, как этого следовало бы ожидать при полной рекапитуляции.
Также было хорошо известно, что, вопреки «закону зародышевого сходства» и «биогенетическому закону», весьма резко различаются по строению самые ранние стадии развития зародышей позвоночных — бластула и гаструла — и лишь на более поздних стадиях развития наблюдается «узел сходства» — стадия, на которой закладывается план строения, характерный для позвоночных, и зародыши всех классов действительно похожи друг на друга. Различия ранних стадий связаны с разным количеством желтка в яйцеклетках: при его увеличении дробление становится сначала неравномерным, а затем (у рыб, птиц и рептилий) неполным поверхностным. В результате меняется и строение бластулы — целобластула имеется у видов с маленьким количеством желтка, амфибластула — со средним и дискобластула — с большим. Кроме того, ход развития на ранних стадиях резко изменяется у наземных позвоночных в связи с появлением зародышевых оболочек.
Связь биогенетического закона с дарвинизмом
Например, если вид А3 возник путём эволюции из более древнего вида А1 через ряд переходных форм (A1 =>А2 => A3), то, в соответствии с биогенетическим законом (в его модифицированном варианте), возможен и обратный процесс, при котором вид А3 превращается в А2 путем укорочения развития и выпадения его конечных стадий (неотения или педогенез).
Дарвинизм и синтетическая теория эволюции, напротив, отрицают возможность полного возврата к предковым формам (Закон необратимости эволюции Долло). Причиной этого, в частности, являются перестройки эмбрионального развития на его ранних стадиях (архаллаксисы по А. Н. Северцову), при которых генетические программы развития меняются настолько существенно, что их полное восстановление в ходе дальнейшей эволюции становится практически невероятным.
Научная критика биогенетического закона и дальнейшее развитие учения о связи онтогенеза и филогенеза
Накопление фактов и теоретические разработки показали, что биогенетический закон в формулировке Геккеля в чистом виде никогда не выполняется. Рекапитуляция может быть только частичной.
Эти факты заставили многих эмбриологов полностью отвергать биогенетической закон в Геккелевских формулировках. Так, С. Гилберт пишет: «Такая точка зрения (о повторении онтогенезом филогенеза) была научно дискредитирована даже раньше, чем была предложена,… Поэтому она распространилась в биологии и общественных науках… прежде, чем было показано, что в ее основе лежат ложные предпосылки»
Столь же резко высказываются Р. Рэфф и Т. Кофмен[12]:»Вторичное открытие и развитие Менделевской генетики на рубеже двух столетий покажет, что в сущности биогенетический закон — это всего лишь иллюзия» (с.30), «Последний удар биогенетическому закону был нанесен тогда, когда стало ясно, что …морфологические адаптации имеют важное значение… для всех стадий онтогенеза» (с.31).
В каком-то смысле в биогенетическом законе перепутаны причины и следствия. Филогенез есть последовательность онтогенезов, следовательно, изменения взрослых форм в ходе филогенеза могут основываться только на изменениях онтогенеза. К такому пониманию соотношения онтогенеза и филогенеза пришел, в частности, А. Н. Северцов, который в 1912—1939 гг разработал теорию филэмбриогенезов[13][1]. Согласно Северцову, все эмбриональные и личиночные признаки делятся на ценогенезы и филэмбриогенезы. Термин «ценогенез», предложенный Геккелем, Северцов трактовал иначе; для Геккеля ценогенез (любые новые признаки, искажавшие рекапитуляцию) был противоположностью палингенеза (сохранения в развитии неизменных признаков, имевшихся и у предков). Северцов термином «ценогенез» обозначал признаки, которые служат приспособлениями к эмбриональному или личиночному образу жизни и у взрослых форм не встречаются, так как не могут иметь для них адаптивного значения. К ценогенезам Северцов относил, например, зародышевые оболочки амниот (амнион, хорион, аллантоис), плаценту млекопитающих, яйцевой зуб зародышей птиц и рептилий и др.
Филэмбриогенезы — это такие изменения онтогенеза, которые в ходе эволюции приводят к изменению признаков взрослых особей. Северцов разделил филэмбриогенезы на анаболии, девиации и архаллаксисы. Анаболия — удлинение онтогенеза, сопровождающееся надставкой стадий. Только при этом способе эволюции наблюдается рекапитуляция — признаки зародышей или личинок потомков напоминают признаки взрослых предков. При девиации происходят изменения на средних стадиях развития, что приводят к более резким изменениям в строении взрослого организма, чем при анаболии. При этом способе эволюции онтогенеза рекапитулировать признаки предковых форм могут лишь ранние стадии потомков. При архаллаксисах изменения происходят на самых ранних стадиях онтогенеза, изменения в строении взрослого организма наиболее часто существенны, а рекапитуляции невозможны.
Литература
- Дарвин Ч., Происхождение видов…, Соч., т. 3, М., 1939
- Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.— Л., 1940
- Козо-Полянский Б. М., Основной биогенетический закон с ботанической точки зрения, Воронеж, 1937
- Северцов А. Н., Морфологические закономерности эволюции, М.—Л., 1939
- Шмальгаузен И. И., Организм как целое в индивидуальном и историческом развитии, М.—Л., 1942
- Мирзоян Э. Н., Индивидуальное развитие и эволюция, М., 1963.
Примечания
- ↑ Значение «Биогенетический закон» в Большой советской энциклопедии
- ↑ Gerhard Medicus (1992). «The Inapplicability of the Biogenetic Rule to Behavioral Development» (PDF). Human Development 35 (1): 1–8. ISSN 0018-716X/92/0351/0001-0008. Проверено 2008-04-30.
- ↑ Северцов А. Н., Морфологические закономерности эволюции, М.—Л., 1939
- ↑ Биогенетический закон — Биологический словарь
- ↑ Н. Н. Воронцов. Эрнст Геккель и судьбы учения Дарвина
- ↑ Биогенетический закон Геккеля-Мюллера
- ↑ 1 2 3 А.А Яхонтов. Зоология для учителя. М., Просвещение, 1985
- ↑ Доказательства эволюции: палеонтология, эмбриология
- ↑ Антропогенез Человека разумного
- ↑ И. И. Мечников. Этюды о природе человека
- ↑ Гилберт С. Биология развития. М., Мир, 1993, т.1, с. 146
- ↑ Р. Рэфф, Т. Кофмен. Эмбрионы, гены и эволюция. М., Мир, 1986, с.30-31
- ↑ Н. Н. Иорданский. Эволюция жизни. М.: Академия, 2001. 425 с.
См. также
Ссылки
dic.academic.ru
БИОГЕНЕТИЧЕСКИЙ ЗАКОН • Большая российская энциклопедия
-
В книжной версии
Том 3. Москва, 2005, стр. 486
-
Скопировать библиографическую ссылку:
Авторы: Н. Н. Иорданский
БИОГЕНЕТИ́ЧЕСКИЙ ЗАКО́Н, концепция, объясняющая взаимоотношения индивидуального развития организмов (онтогенеза) и эволюционной истории видов (филогенеза), обоснованная нем. учёными Ф. Мюллером (1864) и Э. Геккелем (1866). Последний сформулировал осн. положения Б. з.: онтогенез всякого организма представляет краткое и сжатое повторение (рекапитуляцию) филогенеза его предков. По Геккелю, в процессе эволюции появление новых признаков происходит в конце индивидуального развития организмов путём добавления новых стадий, а прежнее взрослое состояние становится предшествующей (последней) эмбриональной стадией. Этим обусловлено повторение предковых состояний в онтогенезе. Т. о., филогенез является причиной формирования онтогенеза.
Индивидуальное развитие не может удлиняться бесконечно, поэтому повторение в эмбриогенезе филогенетич. стадий постепенно становится сокращённым. Как было показано Ч. Дарвином (1859), естественный отбор, воздействуя на все стадии онтогенеза организмов, обусловливает развитие приспособлений каждой из них к особенностям внешних условий. Э. Геккель назвал специфич. приспособления зародышей и личинок ценогенезами. Среди них он различал изменения места формирования эмбриональных зачатков (геторотопии) и сроков и темпов их развития (гетерохронии). Благодаря возникновению ценогенезов в индивидуальном развитии нарушается правильность повторения консервативных признаков и процессов.
Э. Геккель полагал, что в онтогенезе последовательно повторяются целые филогенетич. стадии, благодаря чему можно воссоздавать ход историч. развития вида. Для этой цели он предложил использовать метод тройного параллелизма, основанный на сопоставлении данных эмбриологии, сравнительной анатомии и палеонтологии. Позднее было показано, что благодаря чрезвычайно часто встречающимся ценогенезам полное повторение филогенетич. стадий развития организма в онтогенезе невозможно. Реально наблюдаются рекапитуляции лишь для отд. органов и структур.
А. Н. Северцов в теории филэмбриогенезов показал, что эволюция организмов происходит на основе наследственных изменений любых стадий индивидуального развития; филогенез является рядом последоват. онтогенезов, а Б. з. лишь частный случай соотношений индивидуального и историч. развития организмов, соблюдаемый только для отд. органов при эволюции онтогенеза посредством надставки его последних стадий – анаболии.
bigenc.ru
Биогенетический закон — Википедия
Биогенетический закон Геккеля-Мюллера (также известен под названиями «закон Геккеля», «основной биогенетический закон»): каждое живое существо в своём индивидуальном развитии (онтогенез) повторяет, в известной степени, формы, пройденные его предками или его видом (филогенез)[1].
Зародыши по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию ГеккеляБиогенетический закон сыграл заметную роль в истории развития науки, однако в настоящее время не признается современной биологической наукой[2][3] (см. далее).
История создания
Фактически «биогенетический закон» был сформулирован ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции.
В 1824—1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своём эмбриональном развитии повторяет взрослые формы более примитивных животных[источник не указан 2602 дня].
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своём развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла (он до конца жизни так и не принял эволюционного учения Дарвина), однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» (см. Макроэволюция) и свидетельство происхождения животных одного типа от общего предка.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован (довольно нечётко) английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиночном его состоянии, всех членов одного и того же большого класса» (Дарвин Ч. Соч. М.-Л., 1939, т. 3, с. 636.)[4]
За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер[5]. В своей книге «За Дарвина» (Für Darwin), изданной в 1864 году, он выделяет курсивом мысль: «историческое развитие вида будет отражаться в истории его индивидуального развития».
Краткая афористичная формулировка этого закона была дана в 1866 году немецким естествоиспытателем Эрнстом Геккелем. Краткая формулировка закона звучит следующим образом: Онтогенез есть рекапитуляция филогенеза (во многих переводах — «Онтогенез есть быстрое и краткое повторение филогенеза»)[6].
Факты, противоречащие биогенетическому закону
Уже в XIX веке было известно достаточно фактов, противоречащих биогенетическому закону. Так, были известны многочисленные примеры неотении, при которой в ходе эволюции происходит укорочение онтогенеза и выпадение его конечных стадий. В случае неотении взрослая стадия вида-потомка напоминает личиночную стадию вида-предка, а не наоборот, как этого следовало бы ожидать при полной рекапитуляции.
Также было хорошо известно, что, вопреки «закону зародышевого сходства» и «биогенетическому закону», весьма резко различаются по строению самые ранние стадии развития зародышей позвоночных — бластула и гаструла — и лишь на более поздних стадиях развития наблюдается «узел сходства» — стадия, на которой закладывается план строения, характерный для позвоночных, и зародыши всех классов действительно похожи друг на друга. Различия ранних стадий связаны с разным количеством желтка в яйцеклетках: при его увеличении дробление становится сначала неравномерным, а затем (у рыб, птиц и рептилий) неполным поверхностным. В результате меняется и строение бластулы — целобластула имеется у видов с маленьким количеством желтка, амфибластула — со средним и дискобластула — с большим. Кроме того, ход развития на ранних стадиях резко изменяется у наземных позвоночных в связи с появлением зародышевых оболочек.
Связь биогенетического закона с дарвинизмом
Биогенетический закон часто рассматривается как подтверждение дарвиновской теории эволюции, хотя он вовсе не следует из классического эволюционного учения.
Например, если вид А3 возник путём эволюции из более древнего вида А1 через ряд переходных форм (A1 => А2 => A3), то, в соответствии с биогенетическим законом (в его модифицированном варианте), возможен и обратный процесс, при котором вид А3 превращается в А2 путём укорочения развития и выпадения его конечных стадий (неотения или педогенез).
Дарвинизм и синтетическая теория эволюции отрицают возможность полного возврата к предковым формам (Закон необратимости эволюции Долло). Причиной этого, в частности, являются перестройки эмбрионального развития на его ранних стадиях (архаллаксисы по А. Н. Северцову), при которых генетические программы развития меняются настолько существенно, что их полное восстановление в ходе дальнейшей эволюции становится практически невероятным. Однако в случае неотенической эволюции вида не происходит возврата к предковой форме как таковой, поскольку зародыш определенного биологического вида отражает её лишь приблизительно. Отпадание конечных стадий развития при неотении не отменяет общего возрастания энтропии внутри системы. Таким образом полная картина такого примера эволюции следующая: A1 => А2 => A3 => А4, где А4 в чём-то схож с А2, но не равен ему. Например гусеницы, способные к размножению без перехода ко взрослой стадии, подобны предкам насекомых, но имеют множество морфологических черт, существенно их отличающих.
Научная критика биогенетического закона и дальнейшее развитие учения о связи онтогенеза и филогенеза
Накопление фактов и теоретические разработки показали, что биогенетический закон в формулировке Геккеля в чистом виде никогда не выполняется. Рекапитуляция может быть только частичной.
Эти факты заставили многих эмбриологов полностью отвергать биогенетической закон в Геккелевских формулировках. Так, С. Гилберт пишет: «Такая точка зрения (о повторении онтогенезом филогенеза) была научно дискредитирована даже раньше, чем была предложена,… Поэтому она распространилась в биологии и общественных науках… прежде, чем было показано, что в её основе лежат ложные предпосылки»[7].
Столь же резко высказываются Р. Рэфф и Т. Кофмен[8]: «Вторичное открытие и развитие Менделевской генетики на рубеже двух столетий покажет, что в сущности биогенетический закон — это всего лишь иллюзия» (с.30), «Последний удар биогенетическому закону был нанесен тогда, когда стало ясно, что …морфологические адаптации имеют важное значение… для всех стадий онтогенеза» (с.31).
В каком-то смысле в биогенетическом законе перепутаны причины и следствия. Филогенез есть последовательность онтогенезов, следовательно, изменения взрослых форм в ходе филогенеза могут основываться только на изменениях онтогенеза. К такому пониманию соотношения онтогенеза и филогенеза пришел, в частности, А. Н. Северцов, который в 1912—1939 гг разработал теорию филэмбриогенезов[9][1]. Согласно Северцову, все эмбриональные и личиночные признаки делятся на ценогенезы и филэмбриогенезы. Термин «ценогенез», предложенный Геккелем, Северцов трактовал иначе; для Геккеля ценогенез (любые новые признаки, искажавшие рекапитуляцию) был противоположностью палингенеза (сохранения в развитии неизменных признаков, имевшихся и у предков). Северцов термином «ценогенез» обозначал признаки, которые служат приспособлениями к эмбриональному или личиночному образу жизни и у взрослых форм не встречаются, так как не могут иметь для них адаптивного значения. К ценогенезам Северцов относил, например, зародышевые оболочки амниот (амнион, хорион, аллантоис), плаценту млекопитающих, яйцевой зуб зародышей птиц и рептилий и др.
Филэмбриогенезы — это такие изменения онтогенеза, которые в ходе эволюции приводят к изменению признаков взрослых особей. Северцов разделил филэмбриогенезы на анаболии, девиации и архаллаксисы. Анаболия — удлинение онтогенеза, сопровождающееся надставкой стадий. Только при этом способе эволюции наблюдается рекапитуляция — признаки зародышей или личинок потомков напоминают признаки взрослых предков. При девиации происходят изменения на средних стадиях развития, что приводят к более резким изменениям в строении взрослого организма, чем при анаболии. При этом способе эволюции онтогенеза рекапитулировать признаки предковых форм могут лишь ранние стадии потомков. При архаллаксисах изменения происходят на самых ранних стадиях онтогенеза, изменения в строении взрослого организма наиболее часто существенны, а рекапитуляции невозможны.
Примечания
- ↑ Значение «Биогенетический закон» в Большой советской энциклопедии
- ↑ Gerhard Medicus (1992). «The Inapplicability of the Biogenetic Rule to Behavioral Development» (PDF). Human Development 35 (1): 1–8. ISSN 0018-716X/92/0351/0001-0008. Проверено 2008-04-30.
- ↑ Северцов А. Н., Морфологические закономерности эволюции, М.—Л., 1939
- ↑ Н. Н. Воронцов. Эрнст Геккель и судьбы учения Дарвина
- ↑ Биогенетический закон Геккеля-Мюллера
- ↑ Биогенетический закон // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- ↑ Гилберт С. Биология развития. М., Мир, 1993, т.1, с. 146
- ↑ Р. Рэфф, Т. Кофмен. Эмбрионы, гены и эволюция. М., Мир, 1986, с.30-31
- ↑ Н. Н. Иорданский. Эволюция жизни. М.: Академия, 2001. 425 с.
См. также
Литература
- Дарвин Ч., Происхождение видов…, Соч., том 3, М., 1939
- Биогенетический закон / Иорданский Н. Н. // «Банкетная кампания» 1904 — Большой Иргиз. — М. : Большая российская энциклопедия, 2005. — С. 486. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 3). — ISBN 5-85270-331-1.
- Козо-Полянский Б. М. Основной биогенетический закон с ботанической точки зрения, Воронеж, 1937
- Мирзоян Э. Н. Индивидуальное развитие и эволюция, М., 1963.
- Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.— Л., 1940
- Рекапитуляция // Пустырник — Румчерод. — М. : Большая российская энциклопедия, 2015. — С. 358. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 28). — ISBN 978-5-85270-365-1.
- Северцов А. Н. Морфологические закономерности эволюции, М.—Л., 1939
- Шмальгаузен И. И. Организм как целое в индивидуальном и историческом развитии, М.—Л., 1942
- Биогенетический закон // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
Ссылки
wikipedia.green
Биогенетический закон Википедия
Биогенетический закон Геккеля—Мюллера (также известен под названиями «закон Геккеля», «основной биогенетический закон»): каждое живое существо в своём индивидуальном развитии (онтогенез) повторяет, в известной степени, формы, пройденные его предками или его видом (филогенез)[1].
Зародыши по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию ГеккеляБиогенетический закон сыграл заметную роль в истории развития науки, однако в ХХ веке был опровергнут и не признается современной биологической наукой [2][3] (см. далее).
История создания[ | ]
Фактически «биогенетический закон» был сформулирован ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции.
В 1824—1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своём эмбриональном развитии повторяет взрослые формы более примитивных животных[источник не указан 2991 день].
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своём развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, у
ru-wiki.ru
Биогенетический закон Википедия
Биогенетический закон Геккеля—Мюллера (также известен под названиями «закон Геккеля», «основной биогенетический закон»): каждое живое существо в своём индивидуальном развитии (онтогенез) повторяет, в известной степени, формы, пройденные его предками или его видом (филогенез)[1].
Зародыши по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию ГеккеляБиогенетический закон сыграл заметную роль в истории развития науки, однако в ХХ веке был опровергнут и не признается современной биологической наукой [2][3] (см. далее).
Фактически «биогенетический закон» был сформулирован ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции.
В 1824—1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своём эмбриональном развитии повторяет взрослые формы более примитивных животных[источник не указан 2991 день].
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своём развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла (он до конца жизни так и не принял эволюционного учения Дарвина), однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» (см. Макроэволюция) и свидетельство происхождения животных одного типа от общего предка.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован (довольно нечётко) английским естествоиспытателем
ruwikiorg.ru
Биогенетический закон — Википедия
Биогенетический закон Геккеля-Мюллера (также известен под названиями «закон Геккеля», «основной биогенетический закон»): каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом (филогенез)[1].
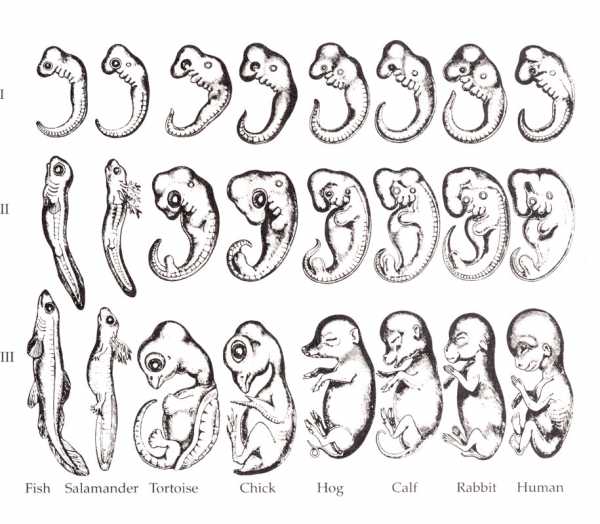 Зародыши по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию Геккеля
Зародыши по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию Геккеля
Геккель был настолько уверен в своём биогенетическом законе, что даже решил подделать доказательства, подкрепляющие его[2][3].
Биогенетический закон сыграл заметную роль в истории развития науки, однако в настоящее время не признается современной биологической наукой[4][5](см. далее ).
История создания[править]
Фактически «биогенетический закон» был сформулирован ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции.
В 1824—1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своем эмбриональном развитии повторяет взрослые формы более примитивных животных[источник не указан 2919 дней].
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своем развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла (он до конца жизни так и не принял эволюционного учения Дарвина), однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» (см. Макроэволюция) и свидетельство происхождения животных одного типа от общего предка.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован (довольно нечётко) английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиночном его состоянии, всех членов одного и того же большого класса» (Дарвин Ч. Соч. М.-Л., 1939, т. 3, с. 636.)[6]
За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер[7]. В своей книге «За Дарвина» (Für Darwin), изданной в 1864 году, он выделяет курсивом мысль: «историческое развитие вида будет отражаться в истории его индивидуального развития».
Краткая афористичная формулировка этого закона была дана в 1866 году немецким естествоиспытателем Эрнстом Геккелем. Краткая формулировка закона звучит следующим образом: Онтогенез есть рекапитуляция филогенеза (во многих переводах — «Онтогенез есть быстрое и краткое повторение филогенеза»)[8].
Факты, противоречащие биогенетическому закону[править]
Уже в XIX веке было известно достаточно фактов, противоречащих биогенетическому закону. Так, были известны многочисленные примеры неотении, при которой в ходе эволюции происходит укорочение онтогенеза и выпадение его конечных стадий. В случае неотении взрослая стадия вида-потомка напоминает личиночную стадию вида-предка, а не наоборот, как этого следовало бы ожидать при полной рекапитуляции.
Также было хорошо известно, что, вопреки «закону зародышевого сходства» и «биогенетическому закону», весьма резко различаются по строению самые ранние стадии развития зародышей позвоночных — бластула и гаструла — и лишь на более поздних стадиях развития наблюдается «узел сходства» — стадия, на которой закладывается план строения, характерный для позвоночных, и зародыши всех классов действительно похожи друг на друга. Различия ранних стадий связаны с разным количеством желтка в яйцеклетках: при его увеличении дробление становится сначала неравномерным, а затем (у рыб, птиц и рептилий) неполным поверхностным. В результате меняется и строение бластулы — целобластула имеется у видов с маленьким количеством желтка, амфибластула — со средним и дискобластула — с большим. Кроме того, ход развития на ранних стадиях резко изменяется у наземных позвоночных в связи с появлением зародышевых оболочек.
Связь биогенетического закона с дарвинизмом[править]
Биогенетический закон часто рассматривается как подтверждение дарвиновской теории эволюции, хотя он вовсе не следует из классического эволюционного учения.
Например, если вид А3 возник путём эволюции из более древнего вида А1 через ряд переходных форм (A1 => А2 => A3), то, в соответствии с биогенетическим законом (в его модифицированном варианте), возможен и обратный процесс, при котором вид А3 превращается в А2 путем укорочения развития и выпадения его конечных стадий (неотения или педогенез).
Дарвинизм и синтетическая теория эволюции отрицают возможность полного возврата к предковым формам (Закон необратимости эволюции Долло). Причиной этого, в частности, являются перестройки эмбрионального развития на его ранних стадиях (архаллаксисы по А. Н. Северцову), при которых генетические программы развития меняются настолько существенно, что их полное восстановление в ходе дальнейшей эволюции становится практически невероятным. Однако в случае неотенической эволюции вида не происходит возврата к предковой форме как таковой, поскольку зародыш определенного биологического вида отражает её лишь приблизительно. Отпадание конечных стадий развития при неотении не отменяет общего возрастания энтропии внутри системы. Таким образом полная картина такого примера эволюции следующая: A1 => А2 => A3 => А4, где А4 в чем-то схож с А2, но не равен ему. Например гусеницы, способные к размножению без перехода ко взрослой стадии, подобны предкам насекомых, но имеют множество морфологических черт, существенно их отличающих.
Научная критика биогенетического закона и дальнейшее развитие учения о связи онтогенеза и филогенеза[править]
Накопление фактов и теоретические разработки показали, что биогенетический закон в формулировке Геккеля в чистом виде никогда не выполняется. Рекапитуляция может быть только частичной.
Эти факты заставили многих эмбриологов полностью отвергать биогенетической закон в Геккелевских формулировках. Так, С. Гилберт пишет: «Такая точка зрения (о повторении онтогенезом филогенеза) была научно дискредитирована даже раньше, чем была предложена,… Поэтому она распространилась в биологии и общественных науках… прежде, чем было показано, что в её основе лежат ложные предпосылки»[9].
Столь же резко высказываются Р. Рэфф и Т. Кофмен[10]: «Вторичное открытие и развитие Менделевской генетики на рубеже двух столетий покажет, что в сущности биогенетический закон — это всего лишь иллюзия» (с.30), «Последний удар биогенетическому закону был нанесен тогда, когда стало ясно, что …морфологические адаптации имеют важное значение… для всех стадий онтогенеза» (с.31).
В каком-то смысле в биогенетическом законе перепутаны причины и следствия. Филогенез есть последовательность онтогенезов, следовательно, изменения взрослых форм в ходе филогенеза могут основываться только на изменениях онтогенеза. К такому пониманию соотношения онтогенеза и филогенеза пришел, в частности, А. Н. Северцов, который в 1912—1939 гг разработал теорию филэмбриогенезов[11][1]. Согласно Северцову, все эмбриональные и личиночные признаки делятся на ценогенезы и филэмбриогенезы. Термин «ценогенез», предложенный Геккелем, Северцов трактовал иначе; для Геккеля ценогенез (любые новые признаки, искажавшие рекапитуляцию) был противоположностью палингенеза (сохранения в развитии неизменных признаков, имевшихся и у предков). Северцов термином «ценогенез» обозначал признаки, которые служат приспособлениями к эмбриональному или личиночному образу жизни и у взрослых форм не встречаются, так как не могут иметь для них адаптивного значения. К ценогенезам Северцов относил, например, зародышевые оболочки амниот (амнион, хорион, аллантоис), плаценту млекопитающих, яйцевой зуб зародышей птиц и рептилий и др.
Филэмбриогенезы — это такие изменения онтогенеза, которые в ходе эволюции приводят к изменению признаков взрослых особей. Северцов разделил филэмбриогенезы на анаболии, девиации и архаллаксисы. Анаболия — удлинение онтогенеза, сопровождающееся надставкой стадий. Только при этом способе эволюции наблюдается рекапитуляция — признаки зародышей или личинок потомков напоминают признаки взрослых предков. При девиации происходят изменения на средних стадиях развития, что приводят к более резким изменениям в строении взрослого организма, чем при анаболии. При этом способе эволюции онтогенеза рекапитулировать признаки предковых форм могут лишь ранние стадии потомков. При архаллаксисах изменения происходят на самых ранних стадиях онтогенеза, изменения в строении взрослого организма наиболее часто существенны, а рекапитуляции невозможны.
- ↑ Значение «Биогенетический закон» в Большой советской энциклопедии
- ↑ Early Evolution and Development: Ernst Haeckel, Evolution 101, University of California Museum of Paleontology, <http://evolution.berkeley.edu/evosite/history/early_evodevo.shtml>. Проверено 20 февраля 2013.
- ↑ Pennisi E. Haeckel’s embryos: fraud rediscovered // Science 1997 277:1435
- ↑ Gerhard Medicus (1992). «The Inapplicability of the Biogenetic Rule to Behavioral Development» (PDF). Human Development 35 (1): 1–8. ISSN 0018-716X/92/0351/0001-0008. Проверено 2008-04-30.
- ↑ Северцов А. Н., Морфологические закономерности эволюции, М.—Л., 1939
- ↑ Н. Н. Воронцов. Эрнст Геккель и судьбы учения Дарвина
- ↑ Биогенетический закон Геккеля-Мюллера
- ↑ Биогенетический закон // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- ↑ Гилберт С. Биология развития. М., Мир, 1993, т.1, с. 146
- ↑ Р. Рэфф, Т. Кофмен. Эмбрионы, гены и эволюция. М., Мир, 1986, с.30-31
- ↑ Н. Н. Иорданский. Эволюция жизни. М.: Академия, 2001. 425 с.
- Дарвин Ч., Происхождение видов…, Соч., том 3, М., 1939
- Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.— Л., 1940
- Козо-Полянский Б. М., Основной биогенетический закон с ботанической точки зрения, Воронеж, 1937
- Северцов А. Н., Морфологические закономерности эволюции, М.—Л., 1939
- Шмальгаузен И. И., Организм как целое в индивидуальном и историческом развитии, М.—Л., 1942
- Мирзоян Э. Н., Индивидуальное развитие и эволюция, М., 1963.
- Биогенетический закон // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
wp.wiki-wiki.ru
история, суть, применение :: SYL.ru
Многие сталкивались с таким понятием, как биогенетические законы развития, но мало кто может объяснить их смысл. Сейчас этот термин употребляется редко из-за критики современных ученых. В чем суть биогенетического закона? Кратко это понятие можно описать так: каждый живой организм в своем развитии проходит в определенной степени те же этапы, что и его предки.

Исторические сведения
Впервые биогенетический закон сформулирован Чарлзом Дарвином в его известном труде «Происхождение видов», изданном в 1859 году. Однако его формулировка была довольно туманной. Более четкое определение понятию биогенетический закон развития дал Эрнст Геккель – знаменитый немецкий ученый, который ввел в обращение такие термины, как онтогенез, экология, филогенез и некоторые другие, а также знаменитый своей теорией о происхождении многоклеточных организмов.
 Формулировка Геккеля гласила, что онтогенез организма является повторением филогенеза, то есть исторического развития организмов того же вида. Биогенетический закон долгое время называли «законом Геккеля», как дань уважения блестящему ученому.
Формулировка Геккеля гласила, что онтогенез организма является повторением филогенеза, то есть исторического развития организмов того же вида. Биогенетический закон долгое время называли «законом Геккеля», как дань уважения блестящему ученому.
Независимо от Геккеля собственное определение биогенетического закона сформулировал другой немецкий естествоиспытатель Фриц Мюллер в 1864 году.
Связь с эволюционной теорией
Модифицированное определение биогенетического закона, согласно которой один вид может приобрести признаки вида, существовавшего ранее, подтверждает эволюционную теорию. Из-за сокращения этапов онтогенеза и архаллаксисов организм приобретает некоторые черты, свойственные его давним предкам, однако приобрести все свойственные им признаки он не может. Это подтверждает второй закон термодинамики для живых организмов (невозможность самопроизвольного уменьшения энтропии в открытой системе) и закон необратимости эволюционных процессов (восстановление утраченных в ходе эволюционного процесса признаков невозможно).
Критика
Биогенетический закон в том виде, в котором его сформулировал Эрнст Геккель, подвергся жесткой критике со стороны исследователей. Большинство ученых сочли доводы коллеги недоказуемыми. Еще в конце 19-го века, когда исследователи захотели узнать, в чем суть биогенетического закона, они обнаружили некоторые противоречия и несоответствия истине. Из наблюдений и экспериментов стало ясно, что онтогенез не полностью, а только отчасти повторяет этапы филогенеза. Примером этого является явление неотении – сокращение онтогенеза и выпадение его отдельных стадий. Неотения характерна для личинок амбистом – аксолотлей, которые из-за индивидуальных гормональных особенностей достигают половой зрелости на стадии личинки.

Понятие, обратное неотении, — анаболия, — определяется как удлинение онтогенеза, появление дополнительных стадий в развитии организма. При такой форме онтогенеза зародыш действительно проходит те же стадии развития, что и его взрослые предки. Однако при анаболии не исключена возможность того, что на поздней стадии развития онтогенез не пойдет другим путем и организм не приобретет определенные отличия от взрослых особей его же вида. То есть, полное повторение всех этапов развития предков того же вида невозможно, так как онтогенез организма происходит под влиянием различных факторов (воздействие окружающей среды, спонтанные мутации в геноме), а не только за счет реализации генетического материала.
Российский биолог А. Северцов ввел термин архаллаксис – такое изменение онтогенеза, при котором самые ранние стадии развития организма отличаются от филогенеза его предков. Очевидно, что рекапитуляция (повторение) признаков, свойственных взрослым особям этого же вида, невозможно, и организм приобретает новые, ранее не свойственные его виду признаки.
Итог
Из ряда научных исследований стало ясно, что сформулированный Геккелем биогенетический закон имеет множество исключений и противоречий. Ученый был уверен в том, что онтогенез полностью повторяет филогенез. В этом была его ошибка. На самом деле филогенез сформирован из ряда онтогенезов представителей определенного вида, а не наоборот. Сейчас термин «биогенетический закон» не применяется в научной литературе.
www.syl.ru