Энграмма — термин, введённый в оборот немецким зоологом и биологом Рихардом Земоном в начале XX века. В теории мнемизма энграмма
Пользователи также искали:
энграмма арк,
энграмма destiny 2,
энграмма физиология,
энграмма простыми словами,
энграмма психофизиология,
энграмма психология,
энграмма тест,
основа энграммы,
энграмма,
Энграмма,
энграмма тест,
энграмма destiny,
энграмма арк,
энграмма физиология,
энграмма простыми словами,
основа энграммы,
энграмма психофизиология,
энграммы,
психофизиология,
тест,
destiny,
физиология,
простыми,
словами,
основа,
психология,
энграмма психология,
энграмма destiny 2,
память.
Общие понятия памяти
Научение может быть рассмотрено как последовательность сложных процессов, вовлекаемых в приобретение, хранение и воспроизведение информации. В результате научения происходит модификация поведения, а память проявляется как сохранение этой модификации.
Наряду с другими познавательными процессами, перцептивными и интеллектуальными, выделяются процессы мнемические (от греч. «мнема» — память).
Мнемические процессы выступают в качестве компонентов познавательной активности человека и неразрывно связаны с его интеллектуальной деятельностью и перцептивными процессами. Образы памяти именуются представлениями.
Память — это процесс сохранения прошлого опыта, делающий возможным его повторное использование в деятельности и возвращение в сферу сознания.
Память связывает прошлое субъекта с его настоящим и оудущим и является познавательной важнейшей функцией, лежащей в основе развития и обучения.
Основой долговременной памяти является формирование энграмм — структурно-функциональных комплексов запоминания информации.
Энграмма — след памяти, сформированный в результате обучения. Описание энграммы может быть выполнено как минимум по трем параметрам:
- динамике развития процессов, приводящих к становлению следа;
- параметру состояния энграммы, характеризующему ее готовность к воспроизведению;
- по устройству энграммы, характеризующему механизмы, которые лежат в основе ее создания.
Эти три разных аспекта описания энграммы составляют основу трех направлений в изучении памяти. Первое исходит из принципа временной организации памяти и описывает динамику формирования энграммы в терминах кратковременного и долговременного хранения; второе, исключая временной компонент создания энграммы, оценивает степень ее готовности к воспроизведению; третье, анализируя нейронные и молекулярные механизмы памяти, может опираться как на принцип временной организации, так и на концепцию состояния энграммы.
Для образования энграмм непременным условием является достаточно длительная реверберация сигналов, связанных с информацией, находящейся в регистре первичной (кратковременной) памяти. Пролонгации реверберации способствуют следующие факторы:
- во-первых, это неоднократное повторение воспринимаемой информации;
- во-вторых, осмысление этой информации, установление её логической структуры или связи с уже хранящейся в долговременной памяти информацией, что резко сокращает количество необходимых повторений;
- в-третьих, установка на длительное запоминание;
- в-четвёртых, высокий интерес к запоминаемому материалу, что даже без повторного восприятия увеличивает длительность реверберации.
Существенное значение имеет функциональное состояние организма, степень утомления, а также эмоциональный фон, связанный с этой информацией.
Beyond Light более наглядное обучение
С релизом дополнения Beyond Light к Destiny 2 в игре изменится вступительный отрезок и обучение, чтобы новичкам было проще освоиться в многопользовательском ролевом шутере.
Когда в прошлом году вышла условно-бесплатная версия Destiny 2, Bungie добавила в игру краткое обучение, чтобы помочь новоприбывшим пользователям. Сперва им нужно было освоиться со стрельбой и снаряжением, а затем победить лёгкого босса. После этих действий открывался доступ к основной игре. Это краткое введение подверглось критике за то, что оно игнорировало основные компоненты геймплея Destiny 2, из-за чего пользователи терялись в большом количестве разнообразных активностей.
Поскольку с выходом Beyond Light практически весь старый контент проекта исчезнет, Bungie решила переработать и начальный этап игры, сделав его более полезным и информативным. Ролик, в котором Призрак находит ваше тело среди руин Космодрома и оживляет вас, останется, но в Destiny 2: Beyond Light будет представлен новый герой: Страж по имени Шоу Хан. Он будет сопровождать игроков в более длительном путешествии, расскажет о состоянии мира, объяснит, что такое энграммы и как перемещаться по многочисленным меню и системам Destiny 2.
Это задание предназначено для новых игроков, но существующие Стражи тоже смогут его пройти после выхода Beyond Light, если захотят.
Кроме того, Bungie подтвердила, что крайний срок переноса учётной записи ПК-версии Destiny 2 из Battle.net в Steam — 1 декабря. Если вы этого не сделаете, то потеряете все сохранения. Дело касается только тех, кто до сих по играет в версию для Battle.net.
Beyond Light выйдет 10 ноября на ПК, PlayStation 4, Xbox One, Xbox Series X и Xbox Series S. На PlayStation 5 расширение станет доступно, когда сама консоль поступит в продажу — 12 ноября (19 ноября в России).
Если вы заметили ошибку — выделите ее мышью и нажмите CTRL+ENTER.
2.6. Основное положение теории активной памяти. Основы психофизиологии
Читайте также
IV. Положение вопроса о зарождении жизни после Пастера
IV. Положение вопроса о зарождении жизни после Пастера
Своими безупречными опытами Пастер, как мы видели, окончательно установил, что произвольного зарождения не происходит. Но такое решение не было утешительным для тех, кто мучился над вопросом о начале жизни: как же
Положение вопроса о зарождении жизни после Пастера
Своими безупречными опытами Пастер, как мы видели, окончательно установил, что произвольного зарождения не происходит. Но такое решение не было утешительным для тех, кто мучился над вопросом о начале жизни: как же
Глава 3 Наша потребность в животных и ее роль для «культурной лестницы». Ручные животные и их значение. Новое положение женщины — снижение ее статуса.
Глава 3 Наша потребность в животных и ее роль для «культурной лестницы». Ручные животные и их значение. Новое положение женщины — снижение ее статуса. Какая же, по моему мнению, черта нашего биологического вида направила его по новому пути и привела к чрезвычайно
Искусственный отбор — основное средство выведения новых пород
Искусственный отбор — основное средство выведения новых пород
Разнообразные превращения видов диких и домашних животных обычно соответствуют меняющимся условиям жизни животных.
Географическое положение Савельевского рудника
Географическое положение Савельевского рудника В 1931 г. была организована добыча горючего сланца в Саратовской области на Савельевском месторождении горючих сланцев, расположенном на правом берегу р. Сакмы (левого притока р. Б. Иргиза) в 35 км к юго-западу от г. Пугачева,
§ 15. Механизмы памяти
§ 15. Механизмы памяти 1. Память — это функция нервных клеток. При синдроме Корсакова, рассеянном склерозе, ишемической болезни мозга, когда дегенерируют нейроны, память исчезает.2. Для способности запоминать надо иметь некоторое количество нейронов, по-видимому, немного
12.
 2. Системы памяти
2. Системы памяти
12.2. Системы памяти Важной вехой в разработке проблем памяти явились работы канадского психолога Д. Хебба (1904–1985). В книге «Организация поведения» он первый дал обоснованную версию механизма образования энграмм как изменения эффективности синапсов (Hebb D., 1949). Эта гипотеза
12.3. Нейрофизиология памяти
12.3. Нейрофизиология памяти Уже в своих основополагающих работах Д. Хебб не обошел вниманием проблему нейрофизиологических механизмов памяти. Что показали дальнейшие наблюдения? Память и научение – это две стороны одной медали. Процессы перевода памяти в долговременную
3.5. Механизмы памяти
3.5. Механизмы памяти
Поведение высших животных было бы невозможно без использования прошлого опыта, т. е. без сохранения информации о нем в нервной системе. Сенсорная память проявляется в удержании в нейроне следа от раздражения рецептора на очень короткое время — до 0,5 с.
е. без сохранения информации о нем в нервной системе. Сенсорная память проявляется в удержании в нейроне следа от раздражения рецептора на очень короткое время — до 0,5 с.
Глава 3 Социальное положение врача в начале XX века и до наших дней, отношения между врачами и пациентами
Глава 3 Социальное положение врача в начале XX века и до наших дней, отношения между врачами и пациентами Примерно до 1940 г. можно было утверждать, что большинство европейского населения проявляло к врачам некоторое уважение, оно сильно зависело от них и без колебания
Положение вирусов в системе живой природы
Положение вирусов в системе живой природы Спорным на протяжении истории биологии является положение в мегасистеме вирусов. Одни авторы считают вирусы особой «неклеточной» формой жизни, другие – что их нельзя относить к живым организмам в полном понимании этого слова,
ТЕОРИИ О ТЕОРИИ ЭВОЛЮЦИИ
ТЕОРИИ О ТЕОРИИ ЭВОЛЮЦИИ
Немало недоразумений возникает из-за неумения отличить общеэволюционный подход от частных метаэволюционных проблем и эти последние друг от друга.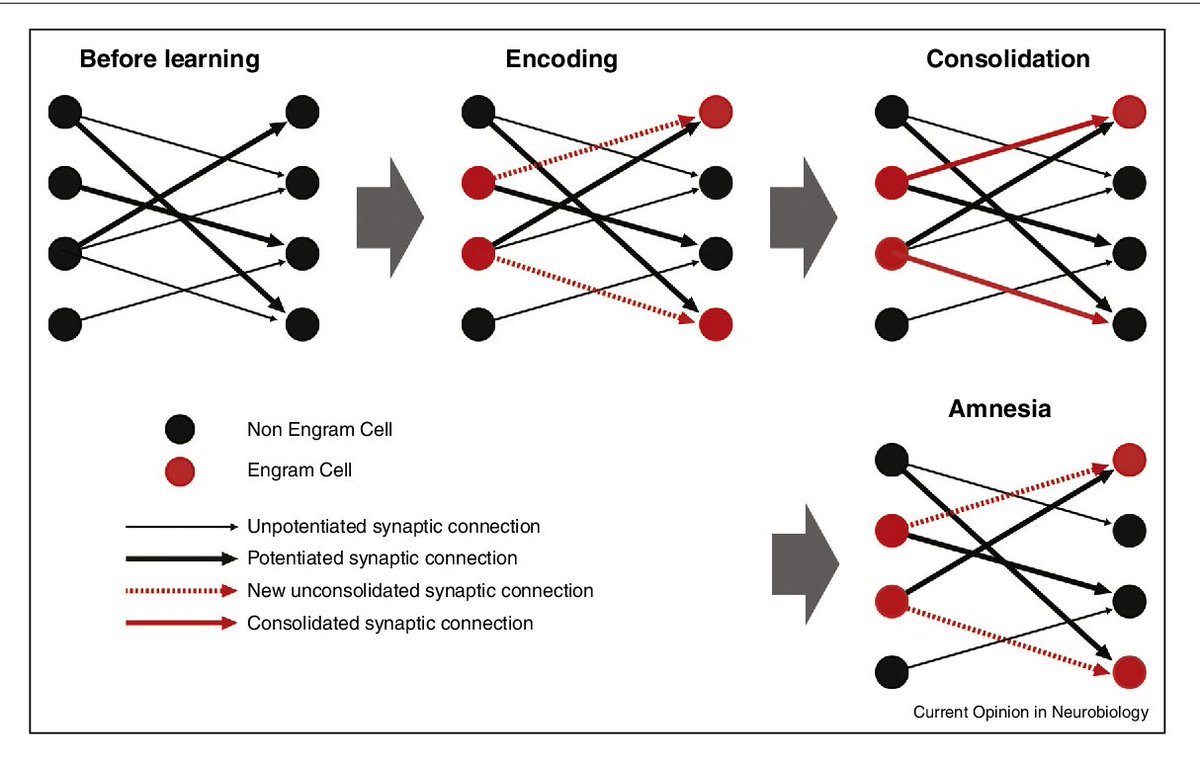 На вопрос, в чем различие между теориями Ж. Б. Ламарка и Ч. Дарвина, большинство отвечает: Ламарк
На вопрос, в чем различие между теориями Ж. Б. Ламарка и Ч. Дарвина, большинство отвечает: Ламарк
18. Положение человека в системе животного мира
18. Положение человека в системе животного мира Вспомните!Назовите общие признаки представителей типа Хордовые; класса Млекопитающие.Данные сравнительной эмбриологии и анатомии человека и других животных позволяют чётко определить, что по критериям зоологической
6.2. Положение и законы термодинамики
6.2. Положение и законы термодинамики Общеизвестно, что термодинамику определяют как науку о законах превращения энергии из одного вида в другой. Существование живых организмов, все разнообразие проявлений жизни также сопровождаются превращением энергии, хотя данная
Лечение голоданием в комбинации с другими методами активной терапии
Лечение голоданием в комбинации с другими методами активной терапии
Во время лечебного голодания могут применяться только бальнеофизиопроцедуры, способствующие процессам выделения и компенсации ацидоза. Эти мероприятия по сути дела входят в комплекс лечения
Эти мероприятия по сути дела входят в комплекс лечения
Destiny 2: Теневые Энграммы
В честь Дня Bungie разработчики добавили кое-что новенькое в Destiny 2, включая ещё один экзотический квест, бонусы в Горниле и Гамбите и новый тип энграмм.
Начался Исход. Нравится вам это или нет, но четыре локации покинут Destiny 2 в следующем сезоне, чтобы освободить место для нового контента. Конечно, Титан, Ио, Марс и Меркурий не могут просто исчезнуть без причины, и эта неделя станет началом конца для некоторых любимых (или не очень) зон патрулирования.
Но из трагических событий можно извлечь пользу: выполняя поручения на планетах, которые скоро будут уничтожены, теперь можно получить энграммы нового типа в качестве награды. Они называются «сфокусированные на исходе Теневые Энграммы» и обладают фантастическим набором лута для фарма, а также дают несколько лёгких триумфов.
Как открыть Теневую Энграмму?
Новые сфокусированные энграммы были представлены в квестовой цепочке «Исход: подготовка». У торговцев на планетах Ио, Марс, Меркурий и Титан появились новые еженедельные поручения. Для прохождения цепочки, вам необходимо выполнить по одному на каждой планете. А в качестве награды за эти поручения вы получите сфокусированные на исходе Теневые Энграммы.
Это новый тип энграмм, который содержит в себе один из четырёх видов оружия:
- гранатомёт «Воспоминания Беренгера»
- снайперскую винтовку «ИКЕЛ_СВ_В1.0.2»
- пистолет-пулемёт «ИКЕЛ_ПП_В1.0.2»
- дробовик «В обратном порядке».
После того, как вы завершите цепочку «Исход: подготовка», расшифровка сфокусированных на исходе Теневых Энграмм будет доступна в Теневом Дешифровщике, который расположен в Башне, недалеко от Скитальца. Обычная расшифровка теневых энграмм откроется автоматически, тогда как улучшенная теневая фокусировка доступна обладателям сезонного пропуска после завершения цепочки в Призмопреобразователе.
Уже после первого его улучшения вы сможете ограничить число предметов, которые выпадают из энграммы, чтобы получить нужный вам лут, превращая обычные теневые энграммы в сфокусированные. Кстати, использовав фокусировку каждого типа по одному разу, вы получите триумф.
Какое оружие Исхода лучше фармить?
К счастью, все четыре вида оружия Исхода относятся к сезону Прибытие, и имеют кап мощи в 1360, а значит, не потеряют актуальности в следующем году. Ранее оружие «ИКЕЛ» было доступно только в новом подземелье «Откровение», не считая оригинальные пушки первого года из эскалационных протоколов, так что теперь у вас есть ещё один способ их получить.
Увы, лучшим оружием «ИКЕЛ» был и остаётся дробовик, а он по-прежнему выбивается только в подземелье.
Впрочем, дробовик «В обратном порядке» – неплохая ему альтернатива, особенно если выпадет с ворпальным перком, великолепно сочетающимся с энергетическими дробовиками. Так что если вы ищете сильное оружие против щитов в PvE или мощный PvP-дробовик, «В обратном порядке» – отличный выбор.
«Воспоминания Беренгара» – это скорострельный гранатомёт, который тоже может выпасть интересными бонусами вроде шанса увеличить боезапас при перезарядке или усиления кинетического урона после взрыва щита, даже если урон оружия не соответствует его типу. С учётом ограничений мощи гранатомётов этого сезона, хорошее оружие этого типа будет в дефиците осенью.
Пока что мы не знаем, какие альтернативы «Воспоминаниям Беренгара» предложит следующий сезон, так что пофармить этот гранатомёт с хорошим роллом кажется неплохой идеей.
Источник
Как получить энграммы в Destiny 2
Автор Admin На чтение 3 мин. Просмотров 4.1k. Опубликовано Обновлено
Destiny 2 – это уникальная, в своем роде, игра, в которой разработчики шагнули далеко вперед, оставив предрассудки и трафареты, которые использовались в других современных играх прежде.
Что такое Энграммы в Destiny 2Например, Энграммы – та самая небольшая частичка Destiny 2, которая и делает игру особенной, и сегодня – мы попробуем разобраться, что это такое и с чем его едят.
Энграммы – это, своего рода смесь, между чертежами, с различных внутриигровых предметов Destiny 2 и сундуками.
Имея определенное количество энграмм – вы сможете получить для себя что-то, начиная от полезной пушки на руки, заканчивая кораблями. Для этого вам нужно будет их расшифровать у криптархов – мастера Рахула или Тиры Карн. Более того – у них вы сможете купить себе еще что-то, что вам понравится.
Какие бывают энграммы в Destiny 2Глобально, все энграммы в игре можно поделить на 5 видов. Это:
Это:
- Обычные энграммы – они не требуют расшифровки, т.к. они не содержат ничего жизненно важного. Обычные предметы, обычное снаряжение.
- Зашифрованные энграммы – а вот эти – уже нужно будет расшифровывать, но опять же – не все. Такие энграммы нередко содержат что-то полезное и редкое.
- Легендарные энграммы – они практически всегда требуют расшифровки криптархами, но и награда, которая в них содержится – крайне полезна.
- Экзотические энграммы – самые ценные в Destiny 2. С них падают просто невероятные устройства.
- Высшие энграммы – также содержат очень крутые вещи.
Понятно, что каждый хранитель хочет получать как можно больше энграмм. И тем более – экзотических. Не стоит забывать и о том, что некоторые экзоты падают только при выполнении определенных миссий и заданий (у нас на сайте вы можете найти несколько таких примеров). Но есть и способы, которые позволят вам фармить крутые энграммы и без дополнительных квестов, и сейчас – мы расскажем вам, как это делается.
Вам понадобится медальон боевой группы. Его вы можете получить с яркой энграммы, которая падает вам при получении очередного уровня. Если нет – вы можете получить там пыль.
Данный медальон – будет стоить 50 единиц пыли.
Если у вас не получается нафармить и пыль – тогда есть мотоциклы и корабли. Вы можете их разбить, и некоторые из них – принесут вам до 100 единиц яркой пыли. Также стоит помнить, что дается медальон не на все время, и действует только 4 часа.
Второе, что вам нужно будет купить – это отчет разведчика, причем именно той планеты, на которой вы находитесь.
С ящиков, которые подсвечиваются – также иногда падают легендарные энграммы. Грубо говоря, на 15 ящиков – вы найдете 1 легендарную энграмму, а в 1 из 30 – экзот. Это вполне стоит того, чтобы побегать – поискать.
Обратите внимание на знамя клана.
Когда он достигнет 3 уровня – в описании знамени вы увидите, что вам будут предлагаться более крутые награды, за открытые мероприятия, а также улучшенные награды, за убийство Кабал.
Вам нужно будет довести открытое мероприятие до героического, т.к. награды там будут еще более крутыми, а шанс на выпадение леги или экзота – увеличится в разы.
В итоге у вас получается: бафф от медальона, расположение сундуков, бафф от клана, героическое мероприятие, а если попадется еще и что-то, связанное с Кабалами – то считайте, что вы счастливчик. В общей сложности – вы сможете нафармить за час – около 2-3 экзотов, и 5-6 лег, правда ведь оно того стоит?
Прочитать позже Отправить статью на e-mail Мы не собираем ваши данные и тем более не передаем их третьим лицам ОтправитьКак получать? / Destiny 2
Энграммы — это предметы (что-то наподобие чертежей), добываемые случайным образом из врагов или получаемые по завершении различных заданий в Destiny 2. В зависимости от типа имеющейся у вас энграммы вы можете получить множество различных предметов — от полезного оружия до кораблей. Тем не менее, для того, чтобы получить предмет снаряжения, энграмму необходимо расшифровать. Для этого отнесите энграмму криптархам: Мастеру Рахулу или Тире Карн, чтобы их расшифровать и раскрыть содержимое.
Тем не менее, для того, чтобы получить предмет снаряжения, энграмму необходимо расшифровать. Для этого отнесите энграмму криптархам: Мастеру Рахулу или Тире Карн, чтобы их расшифровать и раскрыть содержимое.
Существует специальный тип эверверс энграмм, который не расшифровывается криптархами. Они должны быть отнесены Тесс Эверис в Башне. Просто поговорите к ней, и она с радостью откроет различные косметические награды, которые можно получить бесплатно. Вы даже можете купить у нее еще немного, если у вас есть немного серебра – премиальной валюты для микротранзакций в Destiny 2.
Типы энграмм
Стандартные энграммы
| Обычная энграмма Эти энграммы не требуют расшифровки. Когда вы их подбираете, они автоматически открывают обычное снаряжение. |
| Зашифрованная энграмма Энграммы, которые можно расшифровать в редкое снаряжение.  Редкая энграмма с необычными маркировками. Большинство из них автоматически расшифровываются при получении, только некоторые должны быть переданы криптарху для расшифровки. Редкая энграмма с необычными маркировками. Большинство из них автоматически расшифровываются при получении, только некоторые должны быть переданы криптарху для расшифровки. |
| Легендарная энграмма Энграммы, которые можно расшифровать в легендарное снаряжение. Энграмма со сложными маркировками шифрования. Расшифровав ее, криптарх может извлечь особо ценный предмет снаряжения. |
| Экзотическая энграмма Энграммы, которые можно расшифровать в экзотическое снаряжение. Энграмма с необычными маркировками шифрования. Любой криптарх придет в восторг от возможности раскрыть ее подлинную природу. Конечно, многих игроков интересует ответ на вопрос – «Где получить экзотические энграммы?» Вы не можете купить экзотические энграммы. Доступные способы: прохождение миссий, налетов, приключений, публичных мероприятий, рейдов, а такжи матчи в Горнило. Также стоит отметить, что некоторые виды снаряжения (например, Универсальный инструмент MIDA, Буря, Крысиный король, Легенда об Акрии) вы не сможете получить из экзотических энграмм.  Для этого необходимо выполнять соответствующие задания. Для этого необходимо выполнять соответствующие задания. |
| Высшая энграмма Пульсирующая энергией энграмма. Расшифровав ее, криптарх может извлечь особо ценный предмет снаряжения. |
Особые энграммы
| Роковая энграмма Энграмма, судьба которой уже предрешена. В ней содержится отсутствующее у вас экзотическое снаряжение (броня или оружие). |
| Изохронная энграмма Особенная энграмма для тех, кто хочет собрать всю коллекцию экзотического снаряжения. Доступна лишь раз в неделю. Можно приобрести у Зура. |
Энграммы фракций
| Энграмма тактика «Авангарда» Содержит награду за помощь Завале в борьбе с его главными врагами. |
| Энграмма Горнила Содержит награду за успехи в Горниле. |
| Энграмма исследований «Авангарда» Содержит награду за помощь в исследованиях Икоры Рей. |
| Энграмма оружейника Содержит награду за помощь оружейникам. |
| Энграмма ЕМЗ Содержит награду за успехи в Европейской Мертвой зоне. |
| Энграмма Аркологии Содержит награду за успехи в Новотихоокеанской Аркологии на Титане. |
| Энграмма Несса Содержит награду за успехи в Аркадианской долине Несса. |
| Энграмма Ио Содержит награду за успехи на Эхо-плато Ио. |
| Энграмма Маяка Энграмма с подписью ключа, принадлежащего Последователям Осириса.  |
| Энграмма Колыбели Энграмма с собственными маркировками шифрования. Ее наверняка сможет расшифровать кто-нибудь, имеющий доступ к сети Кловиса Брея. |
| Энграмма Новой монархии Содержит награду за успехи, достигнутые во имя «Новой монархии» |
| Энграмма «Культа грядущей войны» Содержит награду за успехи, достигнутые во имя «Культа грядущей войны». |
| Энграмма «Мертвой орбиты» Содержит награду за успехи, достигнутые во имя «Мертвой орбиты». |
| Железная энграмма Содержит награду за успехи в Железном знамени. |
| Имперская энграмма Содержит награду за успехи на борту «Левиафана» императора Калуса.  |
| Энграмма Испытаний Содержит награду за успехи в Испытаниях Девяти. |
Эверверс энграммы
| Яркая энграмма Энграмма с предметами для изменения внешнего вида снаряжения, включающая украшения для экзотических предметов из «Отвергнутых» и расписку на контракт Эверверс (сезон 1). Эту энграмму больше нельзя получить. |
| Светящаяся энграмма Эта яркая энграмма содержит предметы для изменения внешнего вида из второго сезона, включающие в себя украшения и экзотическое снаряжение из расширения «Проклятие Осириса» (сезон 2). Эту энграмму больше нельзя получить. |
| Энграмма-прототип Эта сияющая энграмма содержит предметы для изменения внешнего вида из третьего сезона, включающие в себя украшения для экзотического снаряжения из расширения «Военный разум» (сезон 3).  |
| Прочная энграмма Энграмма с предметами для изменения внешнего вида снаряжения, включающая украшения для экзотических предметов из «Отвергнутых» и расписку на контракт Эверверс (сезон 4). |
| Высеченная энграмма Энграмма с предметами для изменения внешнего вида снаряжения, включающая расписку на контракт Эверверс (сезон 5). |
| Энграмма Рассвета Энграмма с яркими маркировками шифрования. Предметы из «Рассвет» (2017). |
| Багряная энграмма Энграмма с сияющими маркировками шифрования. Предметы из «Багряные дни». |
| Солнцестояние героев Энграмма с сияющими маркировками шифрования. Предметы из «Солнцестояние героев».  |
| Фестиваль усопших Энграмма с сияющими маркировками шифрования. Предметы из «Фестиваль усопших». |
| Заснеженная энграмма Энграмма с сияющими маркировками шифрования. Предметы из «Рассвет» (2018). |
энграмма — обзор | Темы ScienceDirect
14.3 Следы памяти, консолидация и повторная консолидация в соответствии с нейробиологией и потенциальными связями с психоанализом
С точки зрения нейропсихологии, след памяти — это инграмма , гипотетическое средство, с помощью которого информация сохраняется в виде биофизических или биохимических изменений в мозг (и другие нервные ткани) в ответ на внешние раздражители. Остается неясным, какие биофизические или биохимические механизмы лежат в основе или представляют собой инграмму.Однако множество экспериментальных данных, полученных от людей и подопытных животных за последнее столетие, продемонстрировало, что биологические изменения действительно происходят в мозге после нового обучения; эти изменения необходимы для сохранения информации во времени или, другими словами, для хранения воспоминаний.
Электрофизиологические исследования показывают, что вновь усвоенная информация кодируется в мозге в виде паттернов нейрональной активности (Eichenbaum, 2004). Со временем эта информация трансформируется в более устойчивые модификации, которые, кажется, укоренились в молекулярных или структурных формах, таких как структурные модификации существующих синапсов или образование новых (синаптическая пластичность).Этот процесс преобразования деятельности, вызванной новым обучением, в стабильные, длительные модификации получил название консолидация памяти (McGaugh, 2000). Важной особенностью процесса консолидации памяти является то, что в течение ограниченного времени после обучения новая трассировка является лабильной, потому что она может быть легко нарушена несколькими типами мешающих событий. Фактически, эксперименты, которые начались примерно в конце 1800-х — начале 1900-х годов, но затем значительно увеличились за последние 50 лет, показали, что если новое воспоминание подвергается таким проблемам, как травма мозга, судороги, повторное обучение В случае какого-либо явления или различных видов фармакологического лечения он исчезает, и тесты на воспоминания в более поздние сроки показывают амнезию. Это было обнаружено во множестве типов воспоминаний и у животных, включая людей (Squire, Stark, & Clark, 2004). Однако со временем память становится все более стабильной, пока она не станет полностью нечувствительной к сбоям или консолидации. В самом деле, если вмешивающийся вызов возникает через некоторое время после того, как память сформирована, никакого эффекта не наблюдается, и память сохраняется отлично. Следовательно, есть возможность разрушить вновь сформированные воспоминания сразу после их формирования и в течение ограниченного времени.Как долго длится это временное окно возможностей? Ответ на этот вопрос все еще обсуждается. Общие мешающие события, такие как травмы или поражения головного мозга, предполагают, что консолидация памяти занимает относительно много времени, которое, хотя и варьируется в разных воспоминаниях, у людей может занять несколько лет. С другой стороны, фармакологические и молекулярные вмешательства, такие как острая блокада синтеза белка de novo , нарушают воспоминания, только если применяются вскоре после тренировки, но они неэффективны через несколько часов или дней спустя.
Это было обнаружено во множестве типов воспоминаний и у животных, включая людей (Squire, Stark, & Clark, 2004). Однако со временем память становится все более стабильной, пока она не станет полностью нечувствительной к сбоям или консолидации. В самом деле, если вмешивающийся вызов возникает через некоторое время после того, как память сформирована, никакого эффекта не наблюдается, и память сохраняется отлично. Следовательно, есть возможность разрушить вновь сформированные воспоминания сразу после их формирования и в течение ограниченного времени.Как долго длится это временное окно возможностей? Ответ на этот вопрос все еще обсуждается. Общие мешающие события, такие как травмы или поражения головного мозга, предполагают, что консолидация памяти занимает относительно много времени, которое, хотя и варьируется в разных воспоминаниях, у людей может занять несколько лет. С другой стороны, фармакологические и молекулярные вмешательства, такие как острая блокада синтеза белка de novo , нарушают воспоминания, только если применяются вскоре после тренировки, но они неэффективны через несколько часов или дней спустя. Эта временная дихотомия, по-видимому, связана с разными фазами общего процесса консолидации (Alberini, 2011; см. Также главу 5).
Эта временная дихотомия, по-видимому, связана с разными фазами общего процесса консолидации (Alberini, 2011; см. Также главу 5).
Однако ряд сравнительно недавних исследований, расширяющих предыдущие наблюдения, впервые опубликованные в 1960-х годах, показали, что консолидация памяти не основана на уникальном единственном процессе молекулярной консолидации, и что после стабилизации в отношении этих помех воспоминания могут снова вернуться к лабильное состояние в течение ограниченного периода времени при восстановлении или повторной активации.Эти реактивированные воспоминания со временем снова становятся стабильными и нечувствительными к разрушению — процесс, который будет подробно описан позже и получил название , реконсолидация (Alberini, 2005; Alberini, Milekic, & Tronel, 2006; Dudai, 2004; Nader, 2003; Сара, 2000). В главах этой книги резюмируются исследования и обсуждаемые вопросы, которые еще предстоит решить в этой увлекательной области.
Эти открытия по реконсолидации памяти произвели революцию в наших представлениях о формировании, хранении, отзыве и стабильности долговременной памяти или, на самом деле, нестабильной, динамической природе следов памяти. Знание того, что воспоминания после извлечения хрупки, изменчивы и разрушимы, важно по многим причинам. Например, помимо лучшего понимания психических процессов, эти знания дают возможность разработать более точные терапевтические протоколы в области психического здоровья, включая психоанализ и психотерапию, которые конкретно нацелены на внутренние особенности и механизмы мнемонических процессов.
Знание того, что воспоминания после извлечения хрупки, изменчивы и разрушимы, важно по многим причинам. Например, помимо лучшего понимания психических процессов, эти знания дают возможность разработать более точные терапевтические протоколы в области психического здоровья, включая психоанализ и психотерапию, которые конкретно нацелены на внутренние особенности и механизмы мнемонических процессов.
После повторного открытия реконсолидации памяти было продолжено несколько исследований, посвященных изучению эффекта от использования поведенческих или сочетания поведенческих и фармакологических методов для лечения психопатологий, таких как посттравматическое стрессовое расстройство и зависимость (Suris, Smith, Powell, & North, 2012; см. Главы 5, 10, 12 и 13 Глава 5 Глава 10 Глава 12 Глава 13).
Наша цель — обсудить роль реактивации следов в психоанализе. В частности, мы подробно рассмотрим, как следовая реактивация важна в психоаналитическом лечении и как она может представлять модель для объяснения формирования и выражения бессознательных процессов, таких как те, которые характеризуют ФУ.
В 1914 году Фрейд опубликовал «Воспоминание, повторение и проработка» , в котором четко изложена его позиция в отношении аналитической техники, а именно, что катартический метод уступил место ассоциативному методу.Фрейд подчеркивал, что лечение должно включать настоящую психическую работу для пациента, для которого пассивный гипноз больше не является клинически эффективным. Цель состоит в том, чтобы запомнить и «заполнить пробелы в памяти», как утверждает Фрейд, и «преодолеть сопротивление, вызванное вытеснением» (Freud, 1914/1958, p. 148).
Какие механизмы используются, когда пациент в психоаналитической обстановке проходит через процесс вспоминания и проработки? Очевидно, что работа аналитика состоит в том, чтобы облегчить повторное пробуждение или реактивацию воспоминаний и способствовать проработке и заполнению пробелов.Однако вспоминание, как сказал Фрейд и как мы теперь можем видеть в нейробиологических терминах, — это непростой процесс.
Во время психоанализа субъект предпринимает процесс осознания (или осознания) основных источников своего бессознательного поведения, как интеллектуально, так и эмоционально, путем их повторного переживания и перенаправления эмоций на аналитика, а затем их повторной обработки. в новом режиме. Таким образом, запоминание и обработка прошлых воспоминаний в новой аналитической обстановке является ключевым компонентом психоаналитического процесса, независимо от того, используется ли он для изучения разума и его функционирования или для облегчения нарушений в терапевтических процессах.Но как это происходит? Почему повторное переживание эмоций и вспоминание прошлого в новой, настоящей обстановке под руководством аналитика позволяет субъекту распознать свои бессознательные модели поведения и в конечном итоге изменить их, чтобы лучше справляться с реалиями взрослой жизни? Ответ на этот вопрос важен и сложен, потому что он многогранен.
в новом режиме. Таким образом, запоминание и обработка прошлых воспоминаний в новой аналитической обстановке является ключевым компонентом психоаналитического процесса, независимо от того, используется ли он для изучения разума и его функционирования или для облегчения нарушений в терапевтических процессах.Но как это происходит? Почему повторное переживание эмоций и вспоминание прошлого в новой, настоящей обстановке под руководством аналитика позволяет субъекту распознать свои бессознательные модели поведения и в конечном итоге изменить их, чтобы лучше справляться с реалиями взрослой жизни? Ответ на этот вопрос важен и сложен, потому что он многогранен.
Согласно нейробиологическим знаниям и, в частности, появившемуся мнению о том, что нормальные воспоминания существуют в очень динамичном состоянии, мы можем предположить, что психоаналитическая терапия и проработка критически подразумевают новое кодирование, объединение новых следов и повторное объединение извлеченных воспоминаний, все из которых предоставит инструменты для заполнения пробелов и эмоциональной перестройки и перенаправления личного опыта.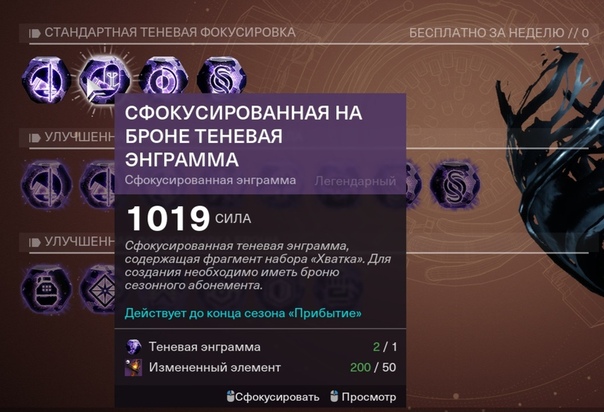 В частности, с повторным открытием реконсолидации памяти большой энтузиазм был направлен на этот процесс памяти, и основные роли реконсолидации были выдвинуты в психоанализе (Bleichmar, 2010; Gorman & Roose, 2011). Мы отмечаем здесь, в целях обсуждения, что остается спорным вопрос о том, является ли реконсолидация механизмом обновления памяти помимо добавления к тому же опыту (см. Tronel et al. (2005) и главу 5). Следовательно, хотя открытие реконсолидации критически изменило наше представление о динамической природе формирования и хранения долговременной памяти, до сих пор неясно, играет ли именно реконсолидация главную роль в динамической перезаписи следов памяти или вместо этого она это формирование новых воспоминаний, которые, следовательно, претерпевают новые процессы консолидации и, следовательно, существуют параллельно со старыми воспоминаниями.Более того, как показывают исследования по повторной консолидации на животных и людях (см. Главы 5 и 10, главу 5, главу 10), повторное объединение декларативных воспоминаний может быть ограничено временем, таким образом затрагивая только недавние, но еще не консолидированные воспоминания.
В частности, с повторным открытием реконсолидации памяти большой энтузиазм был направлен на этот процесс памяти, и основные роли реконсолидации были выдвинуты в психоанализе (Bleichmar, 2010; Gorman & Roose, 2011). Мы отмечаем здесь, в целях обсуждения, что остается спорным вопрос о том, является ли реконсолидация механизмом обновления памяти помимо добавления к тому же опыту (см. Tronel et al. (2005) и главу 5). Следовательно, хотя открытие реконсолидации критически изменило наше представление о динамической природе формирования и хранения долговременной памяти, до сих пор неясно, играет ли именно реконсолидация главную роль в динамической перезаписи следов памяти или вместо этого она это формирование новых воспоминаний, которые, следовательно, претерпевают новые процессы консолидации и, следовательно, существуют параллельно со старыми воспоминаниями.Более того, как показывают исследования по повторной консолидации на животных и людях (см. Главы 5 и 10, главу 5, главу 10), повторное объединение декларативных воспоминаний может быть ограничено временем, таким образом затрагивая только недавние, но еще не консолидированные воспоминания. По сути, это может быть не процесс воссоединения, который в психоаналитическом процессе играет главную роль в обновлении воспоминаний при изменении представления переживаний. Более того, извлечение само по себе не ослабляет и не разрушает воспоминания, а, скорее, может усилить память посредством реконсолидации.В самом деле, если во временном окне хрупкости не происходит никаких помех, память повторно консолидируется и, вероятно, становится сильнее и долговечнее (см. Главы 3, 5 и 10, главу 3, главу 5, главу 10). С другой стороны, мы предполагаем, что новые восприятия, присутствующие в психоаналитическом лечении, и, что важно, новый аффект и эмоциональное состояние настоящего при вспоминании прошлого, действительно предоставляют возможность для изменения консолидированных воспоминаний через новые следы памяти (обновление). или даже, при определенных условиях, ослабление недавних следов в памяти путем вмешательства в их повторное объединение (исчезновение или новое обучение во время повторного объединения; см.
По сути, это может быть не процесс воссоединения, который в психоаналитическом процессе играет главную роль в обновлении воспоминаний при изменении представления переживаний. Более того, извлечение само по себе не ослабляет и не разрушает воспоминания, а, скорее, может усилить память посредством реконсолидации.В самом деле, если во временном окне хрупкости не происходит никаких помех, память повторно консолидируется и, вероятно, становится сильнее и долговечнее (см. Главы 3, 5 и 10, главу 3, главу 5, главу 10). С другой стороны, мы предполагаем, что новые восприятия, присутствующие в психоаналитическом лечении, и, что важно, новый аффект и эмоциональное состояние настоящего при вспоминании прошлого, действительно предоставляют возможность для изменения консолидированных воспоминаний через новые следы памяти (обновление). или даже, при определенных условиях, ослабление недавних следов в памяти путем вмешательства в их повторное объединение (исчезновение или новое обучение во время повторного объединения; см. главы 8 и 9, глава 8, глава 9).
главы 8 и 9, глава 8, глава 9).
Также возможно, как обсуждалось в главе 5, что реконсолидация эмоциональных воспоминаний, которые имеют скорее имплицитную, чем декларативную природу, не могут (или могут быть в меньшей степени) ограничены возрастом памяти. Если это так (что еще нужно доказать), то реконсолидация вместе с консолидацией новых следов может играть более важную роль в психоаналитических условиях. Это понимание будет важно, потому что мы еще не знаем, являются ли следы бессознательной памяти более механистически похожими на имплицитные или явные воспоминания или они следуют другим правилам.В том же духе следует рассмотреть еще один очень важный вопрос, который имеет отношение к обсуждению роли стадий и процессов памяти в психоанализе: следуют ли воспоминания, сформированные в процессе развития, тем же или другим правилам, что и те, которые сформировались в зрелом возрасте. . Поскольку большинство исследований механизмов консолидации и повторной консолидации памяти проводилось на взрослых, важно задаться вопросом, применимы ли те же знания во время развития. Очень необходимы исследования консолидации и обратного уплотнения на этапах разработки.
Очень необходимы исследования консолидации и обратного уплотнения на этапах разработки.
Несмотря на все эти вопросы, которые еще предстоит решить, мы считаем, что, как упоминалось ранее, объединение новых трассировок вместе с обновлением старых воспоминаний посредством консолидации, а также повторное объединение недавних трассировок может способствовать сложному процессу, который происходит. в психоаналитических условиях. Затем мы предлагаем механистическую гипотезу, которая может объяснить такой процесс.
Определение энграммы на Dictionary.com
[en-gram] SHOW IPA
/ ˈɛn græm / PHONETIC RESPELLING
существительное
предполагаемое кодирование в нервной ткани, которое обеспечивает физическую основу для сохранения памяти; след памяти.
ВОПРОСЫ
ПРОГНОЗИРУЙТЕ СЕБЯ ИМЕЕТ ПРОТИВ. ИМЕЮТ!
Есть ли у вас грамматические навыки, чтобы знать, когда использовать «иметь» или «иметь»? Давайте узнаем это с помощью этой викторины!
Вопрос 1 из 7
Моя бабушка ________ Стена, полная старинных часов с кукушкой.
ДРУГИЕ СЛОВА ИЗ энграммы
эн · грамм · микрофон, прилагательноеСлова рядом с энграммой
engrail, engrail, engracked, engrain, engrained, engrail, engraphia, engrave, гравировальный жук, трюк гравера, гравировкаDictionary.com Несокращенный На основе Несокращенного словаря Random House, © Random House, Inc. 2021
Примеры предложений из Интернета для инграммы
.expandable-content {display: none;}. Css-12x6sdt.expandable.content-extended> .expandable-content {display: block;}]]>-
Как мы показали в нашем сравнении, каждое успешное внушение оставляет сильный след или инграмму в мозгу.
-
Эти два закона вместе представляют собой частично гипотезу (инграмму), а частично наблюдаемый факт.
Изучить Dictionary.com
li {-webkit-flex-based: 49%; — ms-flex-предпочтительный размер: 49%; flex-base : 49%;} @ media only screen и (max-width: 769px) {. Css-2jtp0r> li {-webkit-flex-base: 49%; — ms-flex-предпочтительный-размер: 49%; flex-base : 49%;}} @ экран только мультимедиа и (max-width: 480 пикселей) {. Css-2jtp0r> li {-webkit-flex-base: 100%; — ms-flex-предпочтительный размер: 100%; flex- основа: 100%;}}]]>
Css-2jtp0r> li {-webkit-flex-base: 49%; — ms-flex-предпочтительный-размер: 49%; flex-base : 49%;}} @ экран только мультимедиа и (max-width: 480 пикселей) {. Css-2jtp0r> li {-webkit-flex-base: 100%; — ms-flex-предпочтительный размер: 100%; flex- основа: 100%;}}]]> Британский словарь, определения энграммы
существительное
психол физическая основа индивидуальной памяти в мозгу См. также трассировку памяти
Производные формы инграммы
энграмма или инграмма, прилагательноеслово Происхождение энграммы
C20: от немецкой энграммы, от греческого en- in + граммовая буква
Словарь английского языка Коллинза — полное и несокращенное цифровое издание 2012 г. © William Collins Sons & Co.Ltd. 1979, 1986 © HarperCollins Издатели 1998, 2000, 2003, 2005, 2006, 2007, 2009, 2012
Медицинские определения инграммы
n.
Физическое изменение, которое, как считается, происходит в живой нервной ткани в ответ на стимулы, как объяснение памяти. Нейрограмма
Нейрограмма
Медицинский словарь Стедмана American Heritage® Авторское право © 2002, 2001, 1995 компанией Houghton Mifflin. Опубликовано компанией Houghton Mifflin.
Прочие — это Readingli {-webkit-flex-base: 100%; — ms-flex-предпочтительный размер: 100%; flex-base: 100%;} @ media only screen and (max-width: 769px) {.css-1uttx60> li {-webkit-flex-base: 100%; — ms-flex-предпочтительный-размер: 100%; flex-base: 100%;}} @ media only screen and (max-width: 480px) { .css-1uttx60> li {-webkit-flex-base: 100%; — ms-flex-предпочтительный размер: 100%; flex-base: 100%;}}]]>энграмма (нейропсихология) | Психология вики
Оценка |
Биопсихология |
Сравнительный |
Познавательный |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
Энграммы — это гипотетические средства, с помощью которых следы памяти сохраняются как физические или биохимические изменения в мозгу (и другой нервной ткани) в ответ на внешние раздражители.
Их также иногда считают нейронной сетью или фрагментом памяти, иногда используя аналогию с голограммой, чтобы описать ее действие в свете результатов, показывающих, что память, по-видимому, не локализована в мозгу. Существование инграмм постулируется некоторыми научными теориями для объяснения устойчивости памяти и того, как воспоминания хранятся в мозгу. Существование неврологически определенных энграмм не оспаривается, хотя их точный механизм и местоположение были постоянным объектом исследований в течение многих десятилетий.
Термин инграмма был введен Ричардом Семоном и исследован Антоном Павловым. Карл С. Лэшли, американский бихевиорист, известный своим важным вкладом в изучение обучения и памяти, попытался найти инграмму. Его неспособность найти единственный биологический локус памяти (или «инграммы», как он это называл) подсказала ему, что воспоминания не локализованы в одной части мозга, а широко распространены по всей коре.
Лэшли выступал за распределенные репрезентации в результате своей неспособности найти что-либо вроде локализованной инграммы за годы экспериментов с повреждениями (источник: Ранняя работа, Коннекционизм).
Позже исследователь Ричард Ф. Томпсон искал инграмму памяти в мозжечке, а не в коре головного мозга.
Томпсон и его коллеги использовали классическое кондиционирование реакции века у кроликов в поисках инграммы. Они выдыхали воздух на роговицу глаза и сопоставляли его с тоном. Этот воздушный поток обычно вызывает автоматическое мигание. После ряда испытаний они приучили кроликов моргать, когда они слышат звуковой сигнал, даже если воздушная затяжка больше не применялась.Во время эксперимента они наблюдали за несколькими клетками мозга, чтобы попытаться найти инграмму.
Одной областью мозга, которую они наблюдали, которая, по их мнению, была возможной частью инграммы памяти, была латеральное межпозиционное ядро (LIP), когда химически дезактивировалось, это приводило к тому, что кролики, которые ранее были приучены моргать при прослушивании тона, к действовать так, как если бы обусловливание никогда не происходило; однако, когда они повторно активировали LIP, они снова ответили на тональный сигнал морганием глаза.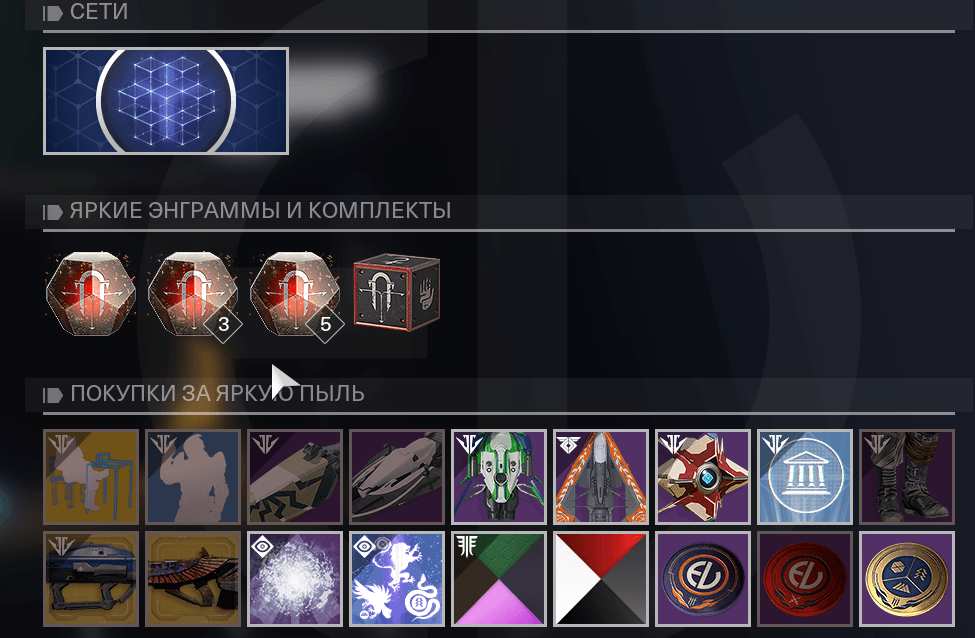 Это свидетельствует о том, что LIP является ключевым элементом инграммы для этой поведенческой реакции. (Джеймс В. Калат, Биологическая психология, стр. 391-393)
Это свидетельствует о том, что LIP является ключевым элементом инграммы для этой поведенческой реакции. (Джеймс В. Калат, Биологическая психология, стр. 391-393)
Важно подчеркнуть, что этот подход, нацеленный на мозжечок, хотя и относительно успешен, исследует только основные, автоматические реакции. Они есть почти у всех животных (особенно в качестве защитных механизмов), и противостоять им довольно сложно. Представьте, что вы пытаетесь не моргнуть, когда кто-то стреляет вам в глаз. В идеале исследование Томпсона и других могло бы в конечном итоге привести к изоляции более сложных инграмм, которые управляют более абстрактными декларативными воспоминаниями, например, как человек помнит свое имя или столицу Франции.
Проблема здесь в том, что значительные исследования показали, что декларативные воспоминания имеют тенденцию перемещаться по мозгу между лимбической системой (глубоко внутри мозга) и внешними областями коры. Это контрастирует с более «примитивным» устройством мозжечка, который контролирует реакцию на моргание и получает прямой ввод слуховой информации. Таким образом, ему не нужно обращаться к другим структурам мозга за помощью в формировании более простых ассоциативных воспоминаний.
Таким образом, ему не нужно обращаться к другим структурам мозга за помощью в формировании более простых ассоциативных воспоминаний.
Инграмма в других контекстах [править | править источник]
Термин инграмма использовался в контекстах, выходящих за рамки нейропсихологии, включая
- Деркс и Холландер (1998) Системное вуду ISBN 18896
- Деркс, Л.И Goldblatt, R., (1985) Концепция сознания с прямой связью: мост между терапевтической практикой и экспериментальной психологией Фонд Уильяма Джеймса, Амстердам.
- Drenth, J.D. (2003) Растущий антиинтеллектуализм в Европе; угроза науке Studia Psychologica, 2003, 45, 5-13
- Sinclair. Дж., Брэй, С. (1992) Азбука НЛП Издатель: Самостоятельно опубликованный (ASPEN) ISBN 0951366017
- Забытые идеи, забытые пионеры: Ричард Семон и история памяти , Даниэль Шактер, 2001
de: Engramm fr: Энграмма
энграмм становятся основной единицей памяти
В новом обзоре Science профессора Сусуму Тонегава из Института обучения и памяти Пикауэра при Массачусетском технологическом институте и Шина Джосселин из больницы для больных детей (SickKids) и Университета Торонто Опишите быстрый прогресс, которого они и коллеги добились за последние десять лет в идентификации, характеристике и даже манипулировании инграммами, а также основные нерешенные вопросы в этой области.
Эксперименты на грызунах показали, что инграммы существуют как многомасштабные сети нейронов. Опыт сохраняется как потенциально извлекаемая память в мозгу, когда возбужденные нейроны в области мозга, такой как гиппокамп или миндалевидное тело, вовлекаются в локальный ансамбль. Эти ансамбли объединяются с другими в других регионах, таких как кора головного мозга, в «комплекс инграмм». Решающее значение для этого процесса связывания инграммных клеток имеет способность нейронов создавать новые связи в цепях посредством процессов, известных как «синаптическая пластичность» и «формирование дендритных шипов».Важно отметить, что эксперименты показывают, что память, изначально сохраненная в комплексе инграммы, может быть восстановлена путем его реактивации, но может также сохраняться «тихо», даже когда воспоминания не могут быть восстановлены естественным образом, например, в моделях мышей, используемых для изучения нарушений памяти, таких как ранняя стадия болезни Альцгеймера. болезнь.
Вверху: клетки энграммы памяти, помеченные зеленым и красным в префронтальной коре головного мозга мыши. Изображение предоставлено: Такаши Китамура
«Более 100 лет назад Семон разработал закон энграфии», — написала Джосселин, старший научный сотрудник SickKids, профессор психологии и физиологии Университета Торонто и старший научный сотрудник программы мозга, разума и сознания Канадского института. for Advanced Research (CIFAR), и Тонегава, профессор биологии и нейробиологии Пикауэра в лаборатории генетики нейронных цепей RIKEN-MIT и исследователь Медицинского института Говарда Хьюза при MIT.«Объединение этих теоретических идей с новыми инструментами, которые позволяют исследователям отображать инграммы и манипулировать ими на уровне клеточных ансамблей, способствовало многим важным открытиям в функции памяти».
«Например, данные указывают на то, что повышенная внутренняя возбудимость и синаптическая пластичность работают рука об руку, чтобы сформировать инграммы, и что эти процессы также могут быть важны для связывания памяти, извлечения памяти и консолидации памяти».
Несмотря на то, что эта область изучила, писали Джосселин и Тонегава, все еще остаются важные неотвеченные вопросы и неиспользованные потенциальные приложения: как инграммы меняются с течением времени? Как можно более непосредственно изучать инграммы и воспоминания на людях? И может ли применение знаний о биологических энграммах вдохновить на развитие искусственного интеллекта, который, в свою очередь, может дать новое понимание работы инграмм?
Статья опубликована в журнале Science , январь.3 издание.
Поиск инграммы | Nature Reviews Neuroscience
Семон, Р. Мнема (Г. Аллен и Анвин, 1921).
Google ученый
Семон Р. Мнемическая психология (Г. Аллен и Анвин, 1923).
Google ученый
Лэшли, К. С. в Симпозиуме Общества экспериментальной биологии, № 4: Психологические механизмы в поведении животных (редакторы Даниелли, Дж. Ф. и Браун, Р.) 454–482 (Academic Press, 1950).
Google ученый
Брюс Д. Пятьдесят лет после работы Лэшли «В поисках энграммы: опровержения и предположения». J. Hist. Neurosci. 10 , 308–318 (2001).
CAS Статья PubMed Google ученый
Дудай Й. и Айзенберг М. Обряды перехода инграммы: реконсолидация и гипотеза затяжной консолидации. Нейрон 44 , 93–100 (2004).
CAS Статья PubMed Google ученый
Джосселин С.А. Продолжение поиска инграммы: исследование механизма воспоминаний о страхе. J. Psychiatry Neurosci. 35 , 221–228 (2010).
PubMed PubMed Central Статья Google ученый
Шактер, Д. L. Забытые идеи, забытые пионеры: Ричард Семон и история памяти (Psychology Press, 2001).
L. Забытые идеи, забытые пионеры: Ричард Семон и история памяти (Psychology Press, 2001).
Google ученый
Москович, М. в Наука памяти: Концепции (ред. Родигер, Х. Л. III, Дудай, Ю. и Фитцпатрик, С. М.) 17–29 (Oxford Univ. Press, 2007).
Google ученый
Кандел, Э. Р. Клеточные основы поведения (W.Х. Фримен, 1976).
Google ученый
Милнер П. М. Автономный мозг: нейронная теория внимания и обучения (Psychology Press, 1999).
Google ученый
Блисс, Т.В. и Коллингридж, Г. Л. Синаптическая модель памяти: долговременная потенциация в гиппокампе. Nature 361 , 31–39 (1993).
CAS Статья Google ученый
Надель, Л. в Наука памяти: концепции (ред. Родигер, Х. Л., Дудай, Ю. и Фицпатрик, С. М.) 177–182 (Oxford Univ. Press, 2007).
Google ученый
Дудай, Ю.Нейробиология. Шаткий след. Nature 406 , 686–687 (2000).
CAS Статья PubMed Google ученый
Надер К. Следы памяти несвязанные. Trends Neurosci. 26 , 65–72 (2003).
CAS Статья PubMed Google ученый
Рамон-и-Кахаль, С. Кронская лекция: тонкая структура нервного центра. Proc. R. Soc. Лондон. 55 , 444–468 (на французском языке) (1894).
Артикул Google ученый
Рамон-и-Кахаль, S. Quelle epoque apparaissent les exsions des cellules nerveuses de la moëlle épinière du poulet? Анат. Anz. 5 , 609–613, 631–639 (на французском языке) (1890).
Google ученый
Stefanowska, M. Les appendices terminaux des dendrites cerebraux et leur different etats Physiologique. Ann. Soc. R. Sci. Med. Nat. Brux. 6 , 351–407 (на французском языке) (1897).
Google ученый
Stefanowska, M. Sur les appendices des dendrites. Бык. Soc. R. Sci. Med. Nat. Brux. 55 , 76–81 (на французском языке) (1897).
Google ученый
Хебб Д. О. Организация поведения: нейропсихологическая теория (Wiley, 1949).
Google ученый
Moser, M. B., Trommald, M. & Andersen, P. Увеличение плотности дендритных шипов на пирамидных клетках CA1 гиппокампа после пространственного обучения у взрослых крыс предполагает образование новых синапсов. Proc. Natl Acad. Sci. США 91 , 12673–12675 (1994).
CAS Статья PubMed Google ученый
Muller, D., Toni, N. & Buchs, P.A. Изменения позвоночника, связанные с длительной потенциацией. Гиппокамп 10 , 596–604 (2000).
CAS Статья PubMed Google ученый
Bosch, M.& Хаяси, Ю. Структурная пластичность дендритных шипов. Curr. Мнение. Neurobiol. 22 , 383–388 (2012).
CAS Статья PubMed Google ученый
Дэй, Дж. Дж. И Свэтт, Дж. Д. Эпигенетические механизмы в познании. Нейрон 70 , 813–829 (2011).
PubMed PubMed Central CAS Статья Google ученый
Сактор Т.C. Как PKMζ поддерживает долговременную память? Нат. Rev. Neurosci. 12 , 9–15 (2011).
CAS Статья PubMed Google ученый
Силва, А.Дж., Коган, Дж. Х., Франкленд, П. В. и Кида, С. CREB и память. Annu. Rev. Neurosci. 21 , 127–148 (1998).
CAS Статья PubMed Google ученый
Джосселин, С. А. и Нгуен, П. В. CREB, синапсы и расстройства памяти: прошлый прогресс и будущие проблемы. Текущее лекарство нацелено на нейрол ЦНС. Disord. 4 , 481–497 (2005).
CAS Статья Google ученый
Хорн, Г. Пути прошлого: отпечаток памяти. Нат. Rev. Neurosci. 5 , 108–120 (2004).
CAS Статья PubMed Google ученый
Рог, Г. Память, импринтинг и мозг: исследование механизмов (Clarendon Press, 1985).
Google ученый
Уитлок, Дж. Р., Хейнен, А. Дж., Шулер, М. Г. и Беар, М. Ф. Обучение вызывает долгосрочное потенцирование в гиппокампе. Наука 313 , 1093–1097 (2006).
Р., Хейнен, А. Дж., Шулер, М. Г. и Беар, М. Ф. Обучение вызывает долгосрочное потенцирование в гиппокампе. Наука 313 , 1093–1097 (2006).
CAS Статья PubMed Google ученый
Румпель, С., LeDoux, J., Zador, A. & Malinow, R. Торговля постсинаптическими рецепторами, лежащая в основе формы ассоциативного обучения. Science 308 , 83–88 (2005).
CAS Статья PubMed Google ученый
Rogan, M. T., Staubli, U. V. & LeDoux, J. E. Обусловленность страхом вызывает ассоциативную долгосрочную потенциацию в миндалине. Nature 390 , 604–607 (1997).
CAS Статья PubMed Google ученый
Oh, M. M., Kuo, A. G., Wu, W. W., Sametsky, E. A. & Disterhoft, J. F. Обучение в водном лабиринте усиливает возбудимость пирамидных нейронов CA1. J. Neurophysiol. 90 , 2171–2179 (2003).
Артикул PubMed Google ученый
Диас, Б. Г. и Ресслер, К. Дж. Обонятельный опыт родителей влияет на поведение и нервную структуру в последующих поколениях. Нат. Neurosci. 17 , 89–96 (2014).
PubMed PubMed Central CAS Статья Google ученый
Графф, Дж. И Мэнсуи, И. М. Эпигенетические коды в познании и поведении. Behav. Brain Res. 192 , 70–87 (2008).
CAS Статья PubMed Google ученый
Barria, A. & Malinow, R. Состав субъединицы рецептора NMDA контролирует синаптическую пластичность путем регулирования связывания с CaMKII. Нейрон 48 , 289–301 (2005).
CAS Статья PubMed Google ученый
Кристенсен, А.S. et al. Механизм регуляции Ca 2+ / кальмодулин-зависимой киназы II стробирования рецептора AMPA..jpg) Нат. Neurosci. 14 , 727–735 (2011).
Нат. Neurosci. 14 , 727–735 (2011).
PubMed PubMed Central CAS Статья Google ученый
Барт, А. Л., Геркин, Р. С. и Дин, К. Л. Изменение свойств возбуждения нейронов после опыта in vivo на трансгенной мыши FosGFP. J. Neurosci. 24 , 6466–6475 (2004).
PubMed PubMed Central CAS Статья Google ученый
Дудай, Ю. в Наука памяти: концепции (ред. Родигер, Х. Л., Дудай, Ю. и Фицпатрик, С. М.) 13–16 (Oxford Univ. Press, 2007).
Google ученый
Уилсон, М.А. и Макнотон, Б. Л. Реактивация ансамблевых воспоминаний гиппокампа во время сна. Science 265 , 676–679 (1994).
Л. Реактивация ансамблевых воспоминаний гиппокампа во время сна. Science 265 , 676–679 (1994).
CAS Статья PubMed Google ученый
Кудримоти, Х.С., Барнс, С.А. и Макнотон, Б.Л. Реактивация ансамблей клеток гиппокампа: эффекты поведенческого состояния, опыта и динамики ЭЭГ. J. Neurosci. 19 , 4090–4101 (1999).
CAS Статья PubMed Google ученый
Надасди З., Хирасе Х., Czurko, A., Csicsvari, J. & Buzsaki, G. Воспроизведение и временная компрессия повторяющихся последовательностей спайков в гиппокампе. J. Neurosci. 19 , 9497–9507 (1999).
CAS Статья PubMed Google ученый
Карр, М. Ф., Джадхав, С. П. и Франк, Л. М. Воспроизведение гиппокампа в состоянии бодрствования: потенциальный субстрат для консолидации и восстановления памяти. Нат. Neurosci. 14 , 147–153 (2011). Исчерпывающий обзор исследований нейронных повторов на грызунах, в которых описывается, как воспроизведение паттернов нейронной активности в бодрствовании и сне способствует восстановлению и консолидации памяти.
PubMed PubMed Central CAS Статья Google ученый
Джи, Д. и Уилсон, М.A. Координированное воспроизведение памяти в зрительной коре и гиппокампе во время сна. Нат. Neurosci. 10 , 100–107 (2007).
CAS Статья PubMed Google ученый
Dupret, D., O’Neill, J., Pleydell-Bouverie, B. & Csicsvari, J. Реорганизация и реактивация карт гиппокампа предсказывают производительность пространственной памяти. Нат. Neurosci. 13 , 995–1002 (2010).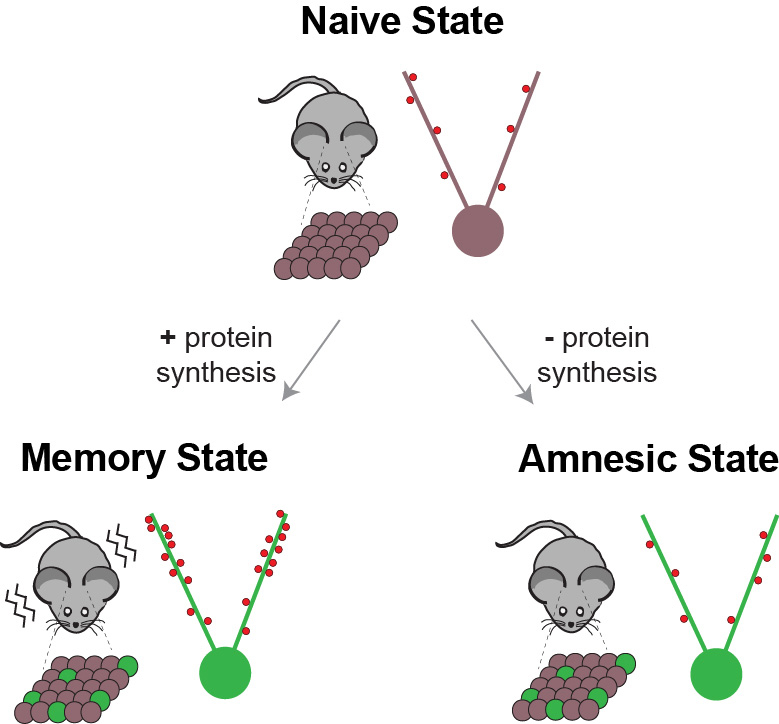
PubMed PubMed Central CAS Статья Google ученый
Накашиба, Т., Buhl, D. L., McHugh, T. J. & Tonegawa, S. Выход CA3 в гиппокампе имеет решающее значение для реактивации, связанной с пульсациями, и консолидации памяти. Нейрон 62 , 781–787 (2009).
PubMed PubMed Central CAS Статья Google ученый
Girardeau, G., Benchenane, K., Wiener, S. I., Buzsaki, G. & Zugaro, M. B. Избирательное подавление гиппокампальной ряби ухудшает пространственную память. Нат.Neurosci. 12 , 1222–1223 (2009).
CAS Статья PubMed Google ученый
Эго-Стенгель, В. и Уилсон, М. А. Нарушение связанной с пульсацией активности гиппокампа во время отдыха ухудшает пространственное обучение у крыс. Гиппокамп 20 , 1–10 (2010).
PubMed PubMed Central Google ученый
Джадхав, С.П. и Франк, Л. М. Восстановление воспоминаний для консолидации. Нейрон 62 , 745–746 (2009).
CAS Статья PubMed Google ученый
Джонсон, А., Ван дер Меер, М. А. и Редиш, А. Д. Интеграция гиппокампа и стриатума в процесс принятия решений. Curr. Мнение. Neurobiol. 17 , 692–697 (2007).
CAS Статья PubMed Google ученый
Штайнер, А.П. и Редиш, А. Д. Поведенческие и нейрофизиологические корреляты сожаления при принятии крысами решения по нейроэкономической задаче. Нат. Neurosci. 17 , 995–1002 (2014).
PubMed PubMed Central CAS Статья Google ученый
Wu, X. & Foster, D. J. Воспроизведение гиппокампа фиксирует уникальную топологическую структуру новой среды. J. Neurosci. 34 , 6459–6469 (2014).
PubMed PubMed Central CAS Статья Google ученый
Джадхав, С. П., Кемере, К., Герман, П. В. и Франк, Л. М. Резкие волны в гиппокампе при пробуждении поддерживают пространственную память. Наука 336 , 1454–1458 (2012).
PubMed PubMed Central CAS Статья Google ученый
Сингер, А. К., Карр, М.Ф., Карлссон, М. П. и Франк, Л. М. Активность КСВ гиппокампа предсказывает правильные решения во время начального обучения задаче чередования. Нейрон 77 , 1163–1173 (2013).
PubMed PubMed Central CAS Статья Google ученый
Карлссон, М. П. и Франк, Л. М. Воспроизведение удаленных переживаний в гиппокампе наяву. Нат. Neurosci. 12 , 913–918 (2009).
PubMed PubMed Central CAS Статья Google ученый
Фостер Д. Дж. И Уилсон М. А. Обратное воспроизведение поведенческих последовательностей в клетках гиппокампа во время бодрствования. Nature 440 , 680–683 (2006).
CAS Статья PubMed Google ученый
Диба, К. и Бузаки, Г. Прямые и обратные последовательности гиппокампа место-клетки во время ряби. Нат. Neurosci. 10 , 1241–1242 (2007).
PubMed PubMed Central CAS Статья Google ученый
Дэвидсон, Т. Дж., Клоостерман, Ф. и Уилсон, М. А. Воспроизведение расширенного опыта в гиппокампе. Нейрон 63 , 497–507 (2009).
PubMed PubMed Central CAS Статья Google ученый
Peigneux, P.и другие. Усиливается ли пространственная память в гиппокампе человека во время медленного сна? Нейрон 44 , 535–545 (2004).
CAS Статья PubMed Google ученый
Maquet, P. et al. Зависящие от опыта изменения церебральной активации во время быстрого сна человека. Нат. Neurosci. 3 , 831–836 (2000).
CAS Статья PubMed Google ученый
Peigneux, P.и другие. Автономное сохранение мозговой активности, связанной с памятью, во время активного бодрствования. PLoS Biol. 4 , e100 (2006).
PubMed PubMed Central Статья CAS Google ученый
Deuker, L. et al. Консолидация памяти путем воспроизведения нейронной активности, специфичной для стимула. J. Neurosci. 33 , 19373–19383 (2013).
PubMed PubMed Central CAS Статья Google ученый
Старесина, Б.П., Алинк, А., Кригескорте, Н. и Хенсон, Р. Н. Реактивация пробуждения предсказывает память у людей. Proc. Natl Acad. Sci. США 110 , 21159–21164 (2013).
CAS Статья PubMed Google ученый
Тамбини, А., Кетц, Н. и Давачи, Л. Усиленные мозговые корреляции во время отдыха связаны с памятью о недавних переживаниях. Нейрон 65 , 280–290 (2010). ФМРТ-исследование человека, показывающее, что паттерны связи гиппокампа и коры головного мозга после задачи ассоциативного кодирования во время отдыха зависят от задачи и предсказывают последующую производительность памяти для этих ассоциаций при извлечении.
PubMed PubMed Central CAS Статья Google ученый
Уотроус, А. Дж., Фелл, Дж., Экстром, А. Д. и Аксмахер, Н. Больше, чем спайки: общие колебательные механизмы для конкретных нейронных репрезентаций во время восприятия и памяти. Curr. Мнение. Neurobiol. 31 , 33–39 (2015).
CAS Статья PubMed Google ученый
Тацуно, М., Lipa, P. & McNaughton, B.L. Методологические соображения по использованию сопоставления шаблонов для изучения длительного воспроизведения трассировки памяти. J. Neurosci. 26 , 10727–10742 (2006).
PubMed PubMed Central CAS Статья Google ученый
Гринберг, М. Э. и Зифф, Э. Б. Стимуляция клеток 3T3 индуцирует транскрипцию протоонкогена c-fos. Nature 311 , 433–438 (1984).
CAS Статья PubMed Google ученый
Курран Т. и Морган Дж. I. Супериндукция c-fos фактором роста нервов в присутствии периферически активных бензодиазепинов. Наука 229 , 1265–1268 (1985).
CAS Статья PubMed Google ученый
Морган, Дж. И. и Карран, Т. Сочетание стимул-транскрипция в нейронах: роль клеточных немедленных генов. Trends Neurosci. 12 , 459–462 (1989).
CAS Статья PubMed Google ученый
Link, W. et al. Соматодендритная экспрессия немедленного раннего гена регулируется синаптической активностью. Proc. Natl Acad. Sci. США 92 , 5734–5738 (1995).
CAS Статья PubMed Google ученый
Лайфорд, Г.L. et al. Arc , фактор роста и ген, регулируемый активностью, кодирует новый ассоциированный с цитоскелетом белок, который обогащен нейрональными дендритами. Нейрон 14 , 433–445 (1995).
CAS Статья PubMed Google ученый
Guzowski, J. F., McNaughton, B. L., Barnes, C. A. & Worley, P. F. Экспрессия немедленного раннего гена Arc в ансамблях нейронов гиппокампа. Нат. Neurosci. 2 , 1120–1124 (1999). Элегантное приложение технологии IEG для идентификации нейронных ансамблей, кодирующих различные контексты.
CAS Статья PubMed Google ученый
Реймерс, Л. Г., Перкинс, Б. Л., Мацуо, Н. и Мэйфорд, М. Локализация стабильного нейронного коррелята ассоциативной памяти. Наука 317 , 1230–1233 (2007). Это исследование представило влиятельный подход TetTag, который позволяет постоянно маркировать нейронные ансамбли, которые активируются во время кодирования памяти страха.Эти авторы показали, что меченые нейроны реактивировались на уровнях выше вероятности, когда позднее вызывается соответствующая память.
CAS Статья PubMed Google ученый
Denny, C.A. et al. Следы памяти гиппокампа по-разному модулируются опытом, временем и взрослым нейрогенезом. Нейрон 83 , 189–201 (2014).
PubMed PubMed Central CAS Статья Google ученый
Guenthner, C.Дж., Миямичи, К., Янг, Х. Х., Хеллер, Х. С. и Луо, Л. Постоянный генетический доступ к временно активным нейронам через TRAP: целенаправленная рекомбинация в активных популяциях. Нейрон 78 , 773–784 (2013).
PubMed PubMed Central CAS Статья Google ученый
Тайлер, К. К., Танака, К. З., Рейджмерс, Л. Г. и Вильтген, Б. Дж. Реактивация нейронных ансамблей во время восстановления недавней и удаленной памяти. Curr. Биол. 23 , 99–106 (2013).
CAS Статья PubMed Google ученый
Лю, X. et al. Оптогенетическая стимуляция инграммы гиппокампа активирует воспоминание о страхе. Природа 484 , 381–385 (2012). Первый пример эксперимента, использующего подход «теги и манипулируй», чтобы убедительно продемонстрировать, что стимуляция инграммы памяти о страхе вызывает искусственное выражение памяти в отсутствие внешних сигналов или внутренних попыток поиска.
PubMed PubMed Central CAS Статья Google ученый
Полин, С. М., Нату, В. С., Коэн, Дж. Д. и Норман, К. А. Специфическая для категории корковая активность предшествует поиску во время поиска в памяти. Наука 310 , 1963–1966 (2005).
CAS Статья PubMed Google ученый
Рагг, М. Д., Джонсон, Дж.Д., Парк, Х. и Ункафер, М. Р. Перекрытие кодирования и поиска в эпизодической памяти человека: функциональная перспектива нейровизуализации. Прог. Brain Res. 169 , 339–352 (2008).
Артикул PubMed Google ученый
Франкленд, П. В. и Бонтемпи, Б. Организация недавних и отдаленных воспоминаний. Нат. Rev. Neurosci. 6 , 119–130 (2005).
CAS Статья PubMed Google ученый
Комияма, Т.и другие. Связанная с обучением мелкомасштабная специфичность, отображаемая в цепях моторной коры у ведущих мышей. Природа 464 , 1182–1186 (2010).
CAS Статья PubMed Google ученый
Ясин Л. и др. Встроенная подсеть высокоактивных нейронов в неокортексе. Нейрон 68 , 1043–1050 (2010).
PubMed PubMed Central CAS Статья Google ученый
МакИнтош, А.Р. К сетевой теории познания. Neural Netw. 13 , 861–870 (2000).
CAS Статья PubMed Google ученый
Maren, S. Нейротоксические поражения базолатеральной миндалины ухудшают обучение и память, но не выполнение условного страха у крыс. J. Neurosci. 19 , 8696–8703 (1999).
CAS Статья PubMed Google ученый
Винокур, Г., Москович, М., Фогель, С., Розенбаум, Р. С. и Секерес, М. Сохранение пространственной памяти после поражений гиппокампа: эффекты обширного опыта в сложной среде. Нат. Neurosci. 8 , 273–275 (2005).
CAS Статья PubMed Google ученый
Сковилл, В. Б. и Милнер, Б. Потеря недавней памяти после двусторонних поражений гиппокампа. J. Neurol. Нейрохирургия. Психиатрия 20 , 11–21 (1957).
PubMed PubMed Central CAS Статья Google ученый
Милнер, Б., Сквайр, Л. Р. и Кандел, Э. Р. Когнитивная нейробиология и исследование памяти. Нейрон 20 , 445–468 (1998).
CAS Статья PubMed Google ученый
О’Киф, Дж. И Достровский, Дж. Гиппокамп как пространственная карта. Предварительные свидетельства активности единиц у свободно передвигающейся крысы. Brain Res. 34 , 171–175 (1971).
CAS Статья PubMed Google ученый
Моррис, Р. Г., Гарруд, П., Роулинз, Дж. Н. и О’Киф, Дж. Навигация по месту у крыс с поражениями гиппокампа. Nature 297 , 681–683 (1982).
CAS Статья PubMed Google ученый
Милнер, П.М. Сотовые сборки: чья идея? Psycholoquy [онлайн], (1999).
Google ученый
Han, J. H. et al. Конкуренция и отбор нейронов при формировании памяти. Наука 316 , 457–460 (2007). Это исследование показывает, что нейроны с более высокой активностью CREB в LA с большей вероятностью, чем их соседи с более низкой активностью CREB, будут отнесены к энграмме памяти о страхе.
CAS Статья PubMed Google ученый
Джосселин, С.A. et al. Долгосрочной памяти способствует сверхэкспрессия белка, связывающего элемент ответа цАМФ, в миндалине. J. Neurosci. 21 , 2404–2412 (2001).
CAS Статья PubMed Google ученый
Дэвис М. Роль миндалевидного тела в страхе и тревоге. Annu. Rev. Neurosci. 15 , 353–375 (1992).
CAS Статья PubMed Google ученый
Марен, С.Миндалевидное тело, синаптическая пластичность и память о страхе. Ann. NY Acad. Sci. 985 , 106–113 (2003).
Артикул PubMed Google ученый
Фанселоу, М. С. и Гейл, Г. Д. Миндалевидное тело, страх и память. Ann. NY Acad. Sci. 985 , 125–134 (2003).
Артикул PubMed Google ученый
Франкленд, П.W. & Josselyn, S.A. Распределение памяти. Нейропсихофармакология 40 , 243 (2015).
Артикул PubMed Google ученый
Zhou, Y. et al. CREB регулирует возбудимость и распределение памяти по подмножествам нейронов миндалевидного тела. Нат. Neurosci. 12 , 1438–1443 (2009).
PubMed PubMed Central CAS Статья Google ученый
Сян, Х.L. et al. Манипулирование «инграммой кокаина» у мышей. J. Neurosci. 34 , 14115–14127 (2014).
PubMed PubMed Central Статья CAS Google ученый
Sano, Y. et al. CREB регулирует распределение памяти в островной коре. Curr. Биол. 24 , 2833–2837 (2014).
PubMed PubMed Central CAS Статья Google ученый
Секерес, М.J. et al. Увеличение функции CRTC1 в зубчатой извилине во время формирования или реактивации памяти увеличивает силу памяти без ущерба для качества памяти. J. Neurosci. 32 , 17857–17868 (2012).
PubMed PubMed Central CAS Статья Google ученый
Yiu, A. P. et al. Нейроны привлекаются к следу памяти, основанному на относительной возбудимости нейронов непосредственно перед тренировкой. Нейрон 83 , 722–735 (2014). Наряду со ссылкой 105, это исследование устанавливает, что размещение нейронов в инграмме регулируется возбудимостью нейронов; то есть более возбудимые клетки с большей вероятностью, чем их менее возбудимые соседи, станут частью инграммы воспоминаний о страхе.
CAS Статья PubMed Google ученый
Армбрустер, Б. Н., Ли, X., Пауш, М. Х., Герлитце, С.И Рот, Б.Л. Разработка замка для соответствия ключу для создания семейства рецепторов, связанных с G-белком, активно активируемых инертным лигандом. Proc. Natl Acad. Sci. США 104 , 5163–5168 (2007).
Артикул CAS PubMed Google ученый
Дейссерот К. Оптогенетика. Нат. Методы 8 , 26–29 (2011).
CAS Статья PubMed Google ученый
Gouty-Colomer, L.A. et al. Выражение Arc идентифицирует боковой след воспоминаний о страхе миндалины. Мол. Психиатрия http://dx.doi.org/10.1038/mp.2015.18 (2015).
Kim, D., Pare, D. & Nair, S. S. Отнесение модельных нейронов миндалины к следу памяти страха зависит от конкурентных синаптических взаимодействий. J. Neurosci. 33 , 14354–14358 (2013).
PubMed PubMed Central CAS Статья Google ученый
Хан, Дж.H. et al. Выборочное стирание воспоминаний о страхе. Наука 323 , 1492–1496 (2009). Используя подход «распределяй и манипулируй», эта статья — первая, показывающая, что воспоминания о страхе можно «стереть» путем удаления инграммных нейронов в ЛП.
CAS Статья PubMed Google ученый
Tanaka, K. Z. et al. Корковые репрезентации восстанавливаются гиппокампом во время восстановления памяти. Нейрон 84 , 347–354 (2014). Недавнее исследование, показывающее, что оптогенетическое молчание нейронов энграммы в гиппокампе предотвращает реактивацию ансамблей инграмм в коре головного мозга, что приводит к неудачному извлечению.
CAS Статья PubMed Google ученый
Koya, E. et al. Целенаправленное разрушение нейронов прилежащего ядра, активируемого кокаином, предотвращает контекстно-зависимую сенсибилизацию. Нат.Neurosci. 12 , 1069–1073 (2009).
PubMed PubMed Central CAS Статья Google ученый
Боссерт, Дж. М. и др. Ансамбли нейронов вентрально-медиальной префронтальной коры опосредуют обусловленный контекстом рецидив героина. Нат. Neurosci. 14 , 420–422 (2011).
PubMed PubMed Central CAS Статья Google ученый
Марр, Д.Простая память: теория архикортекса. Phil. Пер. R. Soc. Лондон. B 262 , 23–81 (1971).
CAS Статья Google ученый
Альварес П. и Сквайр Л. Р. Консолидация памяти и медиальная височная доля: простая сетевая модель. Proc. Natl Acad. Sci. США 91 , 7041–7045 (1994).
CAS Статья PubMed Google ученый
Treves, A.& Роллс, Э. Т. Компьютерный анализ роли гиппокампа в памяти. Гиппокамп 4 , 374–391 (1994).
CAS Статья PubMed Google ученый
van den Heuvel, M. P. & Sporns, O. Сетевые узлы в человеческом мозге. Trends Cogn. Sci. 17 , 683–696 (2013).
Артикул PubMed Google ученый
Уиллер, А.L. et al. Идентификация функционального коннектома для долговременной памяти о страхе у мышей. PLoS Comput. Биол. 9 , e1002853 (2013).
PubMed PubMed Central CAS Статья Google ученый
Penfield, W. & Perot, P. Запись слухового и визуального опыта мозга. Заключительное итоговое обсуждение. Мозг 86 , 595–696 (1963).
CAS Статья PubMed Google ученый
Хамани, К.и другие. Улучшение памяти, вызванное стимуляцией глубокого мозга гипоталамуса / свода. Ann. Neurol. 63 , 119–123 (2008).
Артикул PubMed Google ученый
Hamani, C. et al. Глубокая стимуляция головного мозга переднего ядра таламуса: эффекты электрической стимуляции на судороги, вызванные пилокарпином, и эпилептический статус. Epilepsy Res. 78 , 117–123 (2008).
Артикул PubMed Google ученый
Ван Дж.X. et al. Целенаправленное улучшение кортикально-гиппокампальных сетей мозга и ассоциативной памяти. Наука 345 , 1054–1057 (2014).
PubMed PubMed Central CAS Статья Google ученый
Редондо, Р. Л. и др. Двунаправленное переключение валентности, связанное с инграммой контекстной памяти гиппокампа. Nature 513 , 426–430 (2014).
PubMed PubMed Central CAS Статья Google ученый
Рамирес, С.и другие. Активация инграмм положительной памяти подавляет депрессивное поведение. Nature 522 , 335–339 (2015).
PubMed PubMed Central CAS Статья Google ученый
Гарнер, А. Р. и др. Генерация синтетического следа памяти. Наука 335 , 1513–1516 (2012).
PubMed PubMed Central CAS Статья Google ученый
Cowansage, K.K. et al. Прямая реактивация когерентной неокортикальной памяти контекста. Нейрон 84 , 432–441 (2014). Это исследование показывает, что стимуляции нейронов, связанных с инграммой, в одной области коры (распределенной инграммы) достаточно для восстановления соответствующих воспоминаний о страхе.
PubMed PubMed Central CAS Статья Google ученый
Penfield, W. Энграммы в человеческом мозгу.Механизмы памяти. Proc. R. Soc. Med. 61 , 831–840 (1968).
PubMed PubMed Central CAS Google ученый
Тулвинг, Э. и Томсон, Д. Специфика кодирования и процессы поиска в эпизодической памяти. Psychol. Ред. 80 , 352–373 (1973).
Артикул Google ученый
Ричардс, Б.A. & Frankland, P. W. Конъюнктивный след. Гиппокамп 23 , 207–212 (2013).
Артикул PubMed Google ученый
Бартлетт, Ф. К. Воспоминание: исследование экспериментальной и социальной психологии (Cambridge Univ. Press, 1932).
Google ученый
Шактер Д. Л. Конструктивная память: прошлое и будущее. Dialogues Clin. Neurosci. 14 , 7–18 (2012).
PubMed PubMed Central Google ученый
Ramirez, S. et al. Создание ложных воспоминаний в гиппокампе. Наука 341 , 387–391 (2013). В этом исследовании «ложное» воспоминание было создано путем сочетания искусственной активации нейронов инграммы, кодирующих безопасный (не шокированный) контекст, с ударом стопы.
CAS Статья PubMed Google ученый
Окава, Н.и другие. Искусственное объединение предварительно сохраненной информации для создания качественно новой памяти. Cell Rep. 11 , 261–269 (2015).
CAS Статья PubMed Google ученый
Надер К., Шафе Г. Э. и Леду Дж. Э. Воспоминания о страхе требуют синтеза белка в миндалевидном теле для повторного уплотнения после восстановления. Nature 406 , 722–726 (2000).
CAS Статья PubMed Google ученый
Ким, Дж., Квон, Дж. Т., Ким, Х. С., Джосселин, С. А. и Хан, Дж. Х. Вызов памяти и модификации путем активации нейронов с повышенным CREB. Нат. Neurosci. 17 , 65–72 (2014). Это исследование установило, что искусственно выраженные воспоминания о страхе также подвергаются зависимой от синтеза белку реконсолидации и, следовательно, имеют общие свойства с естественными воспоминаниями о страхе.
CAS Статья PubMed Google ученый
Аллен Т.А., Моррис, А. М., Маттфельд, А. Т., Старк, К. Э. и Фортин, Н. Дж. Модель последовательности событий эпизодической памяти показывает параллели у крыс и людей. Гиппокамп 24 , 1178–1188 (2014).
Артикул PubMed Google ученый
Эйхенбаум, Х. и Фортин, Н. Дж. Преодоление разрыва между мозгом и поведением: когнитивные и нейронные механизмы эпизодической памяти. J. Exp. Анальный. Behav. 84 , 619–629 (2005).
PubMed PubMed Central Статья Google ученый
Тулвинг, Э. Эпизодическая память: от разума к мозгу. Annu. Rev. Psychol. 53 , 1–25 (2002).
Артикул PubMed Google ученый
Лэшли К.С. Интегративные функции коры головного мозга. Physiol. Ред. 13 , 1–42 (1933).
Артикул Google ученый
Лэшли, К.С. Исследования церебральных функций в обучении. XI. Поведение крысы в откидных дверях. Comparative Psychol. Монографии 11 , 5–42 (1935).
Google ученый
Льюис, Д. Дж. Психобиология активной и неактивной памяти. Psychol. Бык. 86 , 1054–1083 (1979).
CAS Статья PubMed Google ученый
Тронель, С., Милекич, М. Х. и Альберини, К. М. Связывание новой информации с реактивированной памятью требует консолидации, а не механизмов повторной консолидации. PLoS Biol. 3 , e293 (2005).
PubMed PubMed Central Статья CAS Google ученый
Дебек, Дж., Дойер, В., Надер, К. и Леду, Дж. Э. Непосредственно реактивированные, но не косвенно реактивируемые воспоминания подвергаются повторной консолидации в миндалевидном теле. Proc. Natl Acad. Sci. США 103 , 3428–3433 (2006).
CAS Статья PubMed Google ученый
О’Киф, Дж. И Надел, Л. Гиппокамп как когнитивная карта (Clarendon Press, 1978).
Google ученый
Олдс, Дж., Дистерхофт, Дж. Ф., Сигал, М., Корнблит, К. Л. и Хирш, Р. Учебные центры мозга крысы, составленные на карту путем измерения латентности реакций условных единиц. J. Neurophysiol. 35 , 202–219 (1972).
CAS Статья PubMed Google ученый
Вайнбергер, Н. М., Бакин, Дж. С. Исследование пластичности слуховой коры. Наука 280 , 1174 (1998).
CAS Статья PubMed Google ученый
Паско, Дж. П. и Капп, Б. С. Электрофизиологические характеристики нейронов центрального ядра миндалины во время выработки условного рефлекса по Павлову у кроликов. Behav. Brain Res. 16 , 117–133 (1985).
CAS Статья PubMed Google ученый
МакКормик, Д. А., Кларк, Г. А., Лавонд, Д. Г. и Томпсон, Р. Ф. Начальная локализация следа памяти для базовой формы обучения. Proc. Natl Acad. Sci. США 79 , 2731–2735 (1982).
CAS Статья PubMed Google ученый
Леду, Дж.Э., Чиккетти, П., Ксагорарис, А. и Романски, Л. М. Боковое миндалевидное ядро: сенсорный интерфейс миндалевидного тела в условиях страха. J. Neurosci. 10 , 1062–1069 (1990).
CAS Статья PubMed Google ученый
Campeau, S. & Davis, M. Вовлечение подкорковых и кортикальных афферентов на латеральное ядро миндалины в кондиционирование страха, измеряемое с помощью потенцированного страхом испуга у крыс, обученных одновременно со слуховыми и визуальными условными стимулами. J. Neurosci. 15 , 2312–2327 (1995).
CAS Статья PubMed Google ученый
Мишкин М. Память у обезьян серьезно нарушена в результате комбинированного, но не раздельного удаления миндалевидного тела и гиппокампа. Nature 273 , 297–298 (1978).
CAS Статья PubMed Google ученый
Коричневый, М.W. & Xiang, J. Z. Память распознавания: нейронные субстраты суждения о предшествующем происшествии. Прог. Neurobiol. 55 , 149–189 (1998).
CAS Статья PubMed Google ученый
Фустер, Дж. М. и Александер, Г. Е. Активность нейронов, связанная с кратковременной памятью. Наука 173 , 652–654 (1971).
CAS Статья PubMed Google ученый
Мияшита, Ю., Камеяма М., Хасегава И. и Фукусима Т. Консолидация визуальной ассоциативной долговременной памяти в височной коре приматов. Neurobiol. Учиться. Mem. 70 , 197–211 (1998).
CAS Статья PubMed Google ученый
Сакаи К. и Мияшита Ю. Нейронная организация долговременной памяти парных партнеров. Nature 354 , 152–155 (1991).
CAS Статья PubMed Google ученый
Рей, Х.G. et al. Записи единичных клеток в медиальной височной доле человека. J. Anat. http://dx.doi.org/10.1111/joa.12228 (2014).
Сквайр, Л. Р. Память и мозг (Oxford Univ. Press, 1987).
Google ученый
Moscovitch, M. et al. Функциональная нейроанатомия удаленной эпизодической, семантической и пространственной памяти: единый учет, основанный на теории множественных следов. J. Anat. 207 , 35–66 (2005).
PubMed PubMed Central Статья Google ученый
Риссман, Дж. И Вагнер, А. Д. Распределенные представления в памяти: выводы из функциональной визуализации мозга. Annu. Rev. Psychol. 63 , 101–128 (2012).
Артикул PubMed Google ученый
Энграммы, появляющиеся как основная единица памяти
Резюме: Визуализация и манипулирование инграммами на клеточном уровне дает важное понимание функции памяти.
Источник: MIT
Хотя ученый Ричард Семон представил концепцию «инграммы» 115 лет назад, чтобы постулировать нейронную основу памяти, прямые свидетельства наличия инграмм начали накапливаться только недавно, поскольку современные технологии и методы начали накапливаться. становятся доступными. В новом обзоре Science профессора Сусуму Тонегава из Института обучения и памяти Пикауэра при Массачусетском технологическом институте и Шина Джосселин из больницы для больных детей (SickKids) и Университета Торонто описывают быстрый прогресс, которого они и коллеги добились последние десять лет в идентификации, характеристике и даже манипулировании инграммами, а также основные нерешенные вопросы в этой области.
Эксперименты на грызунах показали, что инграммы существуют как многомасштабные сети нейронов. Опыт сохраняется как потенциально извлекаемая память в мозгу, когда возбужденные нейроны в области мозга, такой как гиппокамп или миндалевидное тело, вовлекаются в локальный ансамбль. Эти ансамбли объединяются с другими в других регионах, таких как кора головного мозга, в «комплекс инграмм». Решающее значение для этого процесса связывания инграммных клеток имеет способность нейронов создавать новые связи в цепях посредством процессов, известных как «синаптическая пластичность» и «формирование дендритных шипов».Важно отметить, что эксперименты показывают, что память, изначально сохраненная в комплексе инграммы, может быть восстановлена путем его реактивации, но может также сохраняться «тихо», даже когда воспоминания не могут быть восстановлены естественным образом, например, в моделях мышей, используемых для изучения нарушений памяти, таких как ранняя стадия болезни Альцгеймера. болезнь.
«Более 100 лет назад Семон выдвинул закон энграфии», — написала Джосселин, старший научный сотрудник SickKids, профессор психологии и физиологии в Университете Торонто и старший научный сотрудник программы мозга, разума и сознания Канадского института. для перспективных исследований, (CIFAR) и Тонегава, профессор биологии и нейробиологии Пикауэра в лаборатории генетики нейронных цепей RIKEN-MIT в Массачусетском технологическом институте и исследователь Медицинского института Говарда Хьюза.«Объединение этих теоретических идей с новыми инструментами, которые позволяют исследователям отображать инграммы и манипулировать ими на уровне клеточных ансамблей, способствовало многим важным открытиям в функции памяти».
«Например, данные показывают, что как повышенная внутренняя возбудимость, так и синаптическая пластичность работают рука об руку, формируя инграммы, и что эти процессы также могут быть важны для связывания памяти, извлечения памяти и консолидации памяти».
Клетки энграммы памяти, помеченные зеленым и красным в префронтальной коре головного мозга мыши.Изображение предоставлено Такаши Китамура / MIT Picower Institute.Несмотря на то, что эта область изучила, писали Джосселин и Тонегава, все еще остаются важные неотвеченные вопросы и неиспользованные потенциальные приложения: как инграммы меняются с течением времени? Как можно более непосредственно изучать инграммы и воспоминания на людях? И может ли применение знаний о биологических энграммах вдохновить на развитие искусственного интеллекта, который, в свою очередь, может дать новое понимание работы инграмм?
Об этой статье исследования нейробиологии
Источник:
MIT
Контакты для СМИ:
Дэвид Оренштейн — MIT
Источник изображения:
Изображение предоставлено Такаши Китамура / MIT Picower Institute.
См. Также
Оригинальное исследование: Результаты появятся в Science .
Не стесняйтесь поделиться этими новостями нейробиологии.
нейронов энграмм: новый подход к консолидации памяти
Источник: StockSnap / Pixabay
Еще при Платоне и Аристотеле люди считали, что наши воспоминания должны быть физическими вещами, которые хранятся где-то в мозгу. Но только в наше время мы многое узнали о том, что это такое.Сначала чему-то было дано имя: энграмма памяти. Затем, по мере накопления знаний о том, что происходит в нейронах и их синапсах, когда они становятся активными в обучении и запоминании, стало ясно, что обучающие события, которые можно запомнить, вызывают химические и физические изменения в соединениях (синапсах) между нейронами, которые участвуют в опыт обучения.
Участвующие нейроны вырастают новые дендритные ветви (называемые шипами), синапсы на этих шипах увеличиваются, а их системы нейротрансмиттеров усиливаются.Эти изменения составляют инграмму. Реактивация синапсов, содержащих такую инграмму, после обучения, может вызвать воспоминание об исходном обучении, создавшем инграмму.
На заре нейробиологии ученые считали, что процесс обучения назначал или задействовал определенные части мозга для удержания памяти. Экспериментатор Карл Лэшли обучал лабораторных животных определенным задачам, а затем под наркозом разрушал различные части неокортекса в надежде найти, где хранится память.Он не смог найти ни одного конкретного хранилища. Что он действительно обнаружил, так это то, что чем более обширны корковые поражения, тем больше вероятность, что он сможет стереть воспоминания. Другими словами, воспоминания о данном опыте, казалось, были деконструированы и разделены на разные области.
Затем последовали количественные исследования ЭЭГ, проведенные Э. Роем Джоном, в которых он отслеживал местоположение вызванных электричеством ответов мозга в различных частях коры во время обучения. Он увидел, что данный опыт обучения вызовет электрические реакции в нескольких частях коры, что снова предполагает деконструкцию и распределение инграмм памяти.Это привело его к известному заявлению: «Память — это не вещь в месте, а процесс в популяции».
Ну, мы знаем, что это немного преувеличение. Есть такая вещь, как инграмма памяти, которая хранится в определенных местах. Тем не менее, существует процесс распределения для создания инграммы в нескольких местах и для их оркестровки в одновременную и скоординированную деятельность во время вызова памяти.
Современная генная инженерия и технология окрашивания нейронов предоставляют новые мощные инструменты для исследования нейронов, которые участвуют в присоединении к энграммам, задействованным в нейронных цепях.Теперь есть способы отображать инграммы и манипулировать ими на уровне нейронных ансамблей. Несколько линий доказательств показывают, что нейроны инграмм можно увидеть гистологически и оценить с помощью различных экспериментальных подходов.
Например, гистологические пятна, выявляющие нейроны, которые активируются в процессе обучения, показывают, что они также активны во время восстановления памяти этого опыта. Во-вторых, исследования потери функции показывают, что нарушение функции нейрона энграммы после опыта ухудшает последующее восстановление памяти.В-третьих, исследования показывают, что восстановление памяти может запускаться оптогенетической стимуляцией нейронов инграммы в отсутствие каких-либо естественных сенсорных сигналов.
Основной подход, который использовали исследователи в лаборатории Сусуму Тонегава, заключался в том, чтобы научить мышей избегать захода в камеру, в которой они могут получить легкий электрический шок. Нейроны, которые активируются этой обусловливающей страх флуоресценцией в иммуногистологических окрашивателях срезов мозга мышей, которых приносят в жертву в разное время после обучения, обнаруживают энграмму памяти, которая находится в выбранных нейронах миндалины (которая обрабатывает информацию о страхе), в гиппокампе (которая преобразует от кратковременной памяти к долговременной памяти), а также во многих областях неокортекса (которая хранит долговременную память в форме расширенных синаптических возможностей).Некоторые из этих клеток все еще флуоресцируют, если их исследовать много дней спустя, что указывает на то, что они стали частью ансамбля инграммных нейронов, которые сохраняют относительно длительное представление об исходном приобретенном опыте.
Другие мыши были генетически сконструированы таким образом, что клетки инграммы флуоресцируют и активируются при воздействии света, доставляемого через микроволоконные кабели, хирургически имплантированные в различные области неокортекса. Такая световая стимуляция инграммных клеток подтвердила их статус инграммы, потому что только световая стимуляция запускала ранее усвоенное поведение (замораживание на месте, а не попадание в шоковую камеру).Ключевым открытием было то, что нейроны инграмм в префронтальной коре головного мозга «молчали» вскоре после обучения — они могли инициировать замораживание при искусственной активации светом, доставляемым через хирургически имплантированные оптоволоконные волокна, но они не срабатывали при естественном воспроизведении памяти. Другими словами, инграмма памяти была сформирована сразу во всех трех местах (миндалевидное тело, гиппокамп и неокортекс), но клетки инграммы в неокортексе должны были со временем созреть, чтобы стать полностью функциональными.
В течение следующих двух недель нейроны инграмм в неокортексе постепенно созревали, что отражалось в изменениях их анатомии и физиологической активности.К концу того же периода клетки инграммы гиппокампа замолчали и больше не использовались для естественного воспроизведения. На этом этапе мыши могли вспомнить событие естественным образом, без активации неокортикальных клеток оптоволоконным светом. Однако следы воспоминаний остались в гиппокампе, потому что реактивация этих нейронов гиппокампа светом заставляла животных замерзать.
В прошлом преобладало мнение, что опыт обучения временно сохраняется в цепях гиппокампа, а затем экспортируется в другие части мозга для окончательного хранения.Как в прошлом, так и сейчас, все свидетельства указывают на то, что гиппокамп имеет решающее значение для формирования долговременных воспоминаний о переживаниях, которые не связаны с моторным обучением, но механизмы были неопределенными. Нейробиологи знали, что долговременные воспоминания хранились вне гиппокампа, потому что люди с повреждением гиппокампа могут потерять способность формировать новые долговременные воспоминания, но они все еще могут вспоминать старые воспоминания.
Новое исследование предполагает, что энграммы памяти не переносятся из гиппокампа в неокортекс, а присутствуют в обоих местах в начале обучения.Инграмма памяти в неокортексе просто требует созревания, чтобы память стала более постоянной. Более того, гиппокамп не может и не должен удерживать длительные инграммы.
Хотя это новый способ размышления о механизмах объединения временных воспоминаний в более длительные, общепринятая идея объединения остается подтвержденной. То есть инграмма памяти должна со временем созревать в виде биохимических и анатомических изменений в клетках инграммы. Очевидно, что такой процесс созревания будет нарушен, если те же самые клетки инграммы будут задействованы для других целей обучения до того, как они завершат свое созревание в качестве конкретной инграммы памяти.Это также помогает объяснить, почему последующие репетиции помогают продлить воспоминания, потому что каждая репетиция повторно вовлекает нейроны инграммы в тот же вид деятельности, который они выполняли во время обучения, тем самым укрепляя соответствующие синапсы.
После того, как у мышей, обусловленных страхом, сформировались воспоминания, клетки инграммы в миндалевидном теле остались неизменными на протяжении всего эксперимента. Те клетки, которые необходимы для вызова эмоций, связанных с конкретными воспоминаниями, такими как в данном случае страх попасть в шоковую камеру, общаются с клетками инграммы как в гиппокампе, так и в префронтальной коре.
Мы не знаем, что происходит с инграммными клетками, специфичными для памяти, в гиппокампе. Может быть, по мере того, как они постепенно теряют свой статус инграммы, они становятся доступными для обработки новых видов опыта обучения. Возможно, некоторые следы инграммы остаются в гиппокампе и доступны для реактивации, если будут получены очень важные входные данные, как это может быть в случае с сильными сигналами памяти. Возможно, важным моментом является то, что эти новые методы маркировки клеток инграммы открывают двери для новых способов изучения восстановления памяти, аспекта механизмов памяти, которым долгое время не уделялось должного внимания.
Еще одно потенциально важное открытие этого вида исследований состоит в том, что энграммы памяти могут быть повреждены, но все еще могут существовать в форме, которую невозможно восстановить естественными способами. Тот факт, что такие «безмолвные» инграммы могут быть получены с помощью прямой оптогенетической стимуляции, указывает на то, что неспособность вспомнить, не обязательно означает, что память потеряна. Проблема может заключаться в неадекватности естественных сигналов памяти, используемых для запуска извлечения памяти.
Также теперь открыта дверь для экспериментов, которые могут продвинуть наше понимание созревания нейронов энграммы в неокортексе.До сих пор известно, что для созревания требуется начальная коммуникация с клетками инграммы в гиппокампе.