Индивидуальное развитие организмов — онтогенез
Онтогенезом называют совокупность процессов, протекающих в организме, с момента образования зиготы до смерти. Его подразделяют на два этапа: эмбриональный и постэмбриональный.
Эмбриональный период
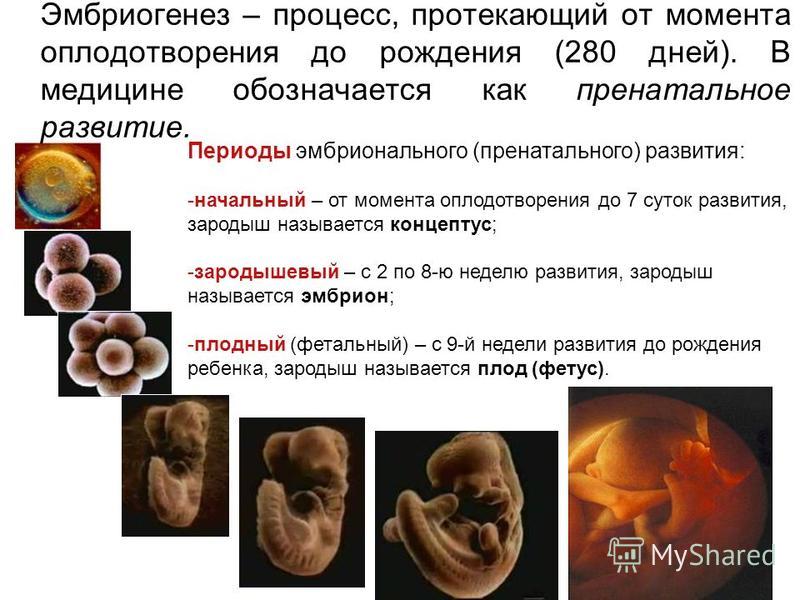
Эмбриональным считают период зародышевого развития с момента образования зиготы до выхода из яйцевых оболочек или рождения, в процессе зародышевого развития эмбрион проходит стадии дробления, гаструляцяя, первичного органогенеза и дальнейшей дифференцировки органов и тканей.
Дробленое. Дроблением называют процесс образования многоклеточного однослойного аародыша — бластулы. Для дробления характерно: 1) деление клеток путем митоза с сохранением диплоидного набора хромосом; 2) очень короткий митотический цикл; 3) бластомеры не дифференцированы, и в них не используется наследственная информация;4) бластомеры не растут и в дальнейшем становятся все меньше; 5) цитоплазма зиготы не перемешивается и не перемещается.
Первая борозда дробления проходит в меридио-нальной ллрркости, соединяющей оба полюса -вегетатив-ный и аяимальный,-и делит зиготу на две одинаковые клетки. Это стадия двух бластомеров. Вторая борозда также меридиональна, но перпендикулярна первой. Она разделяет оба бластомера, возникших в результате первого деления, надвое — образуются четыре сходных бластомера. Следующая, третья, борозда дробления — широтная. Она пролегает несколько выше экватора и делит все четыре бластомера сразу на во-семь клеток. В дальнейшем борозды дробления чередуются. По мере увеличения числа клеток деление их становится не-одновременным. Бластомеры все дальше и дальше отходят от центра зародыша, образуя полость. В конце дробления зародыш принимает форму пузырька со стенкой, образованной одним слоем клеток, тесно прилегающих друг к другу. Внутренняя полость зародыша, первоначально сообщавшаяся с внешней сре-дой через щели между бластомерами, в результате их плотного смыкания становится совершенно изолированной.
Дробление оплодотворенной яйцеклетки может происходить по-разному. Яйцо ланцетника дробится полностью и имеет равные по размеру бластомеры. Такой тип дробления называется
Третий тип дробления характерен для яйцеклеток птиц, рептилий, у которых желтка много, и называется дискоидалъным. Здесь в дробление вовлекается только ядро и тонкий участок цитоплазмы, в результате образуется зародышевый диск (желток яйца при этом не дробится). У яиц членистоногих (желток сосредоточен в центре яйцеклетки) дробление 
При полном дроблении (например, у ланцетника на стадии 32 бластомеров) зародыш имеет вид тутовой ягоды и называется морулой. Приблизительно на стадии 64 бластомеров в нем формируется полость, а бластомеры располагаются в один слой, образуя стенку зародыша. Эта стадия зародыша называется бластулой. Вскоре начинается процесс возникновения двухслойного зародыша —
Существует несколько способов гаструляции. Первый -иммиграция — наблюдается у кишечнополостных: после образования бластулы некоторые клетки стенки тела зародыша иммигрируют в глубь полости и постепенно заполняют ее.
У амфибий гаструла образуется по-другому: более мелкие бластомеры со стороны анимального полюса наползают поверх крупных бластомеров вегетативного полюса, так что двухслойный зародыш получается путем обрастания мелкими бластомерами крупных. У членистоногих бластомеры в ходе дробления отделяют от себя дочерние клетки внутрь полости, где они образуют второй слой зародыша — энтодерму. Этот способ возникновения гаструлы называется расщеплением. Различные способы формирования двухслойного зародыша у разных видов животных обусловлены количеством и характером распределения желтка в яйце. Однако строго обособленных типов гаструляции не наблюдается, их подразделение условно.
Этот способ возникновения гаструлы называется расщеплением. Различные способы формирования двухслойного зародыша у разных видов животных обусловлены количеством и характером распределения желтка в яйце. Однако строго обособленных типов гаструляции не наблюдается, их подразделение условно.
Первичный органогенез. После завершения гаструляции у зародыша образуется комплекс осевых органов: нервная трубка, хорда, кишечная трубка. Начиная с плоских червей в эволюции животного мира наступило крупное усложнение: в зародыше закладывается третий зародышевый листок — мезодерма. У хордовых это происходит путем отшнуровывания от энтодермы мезодермальных карманов, которые врастают между первым и вторым зародышевыми листками, формируя вторичную полость тела.
Дальнейшая дифференцировка клеток зародыша приводит к возникновению многочисленных производных зародыше-вых листков-органов и тканей.
Дифференцировка или дифференцирование — это про-цесс возникновения и нарастания структурных и функцио-нальных различий между отдельными клетками и частями зародыша. С морфологической точки зрения дифференцирование выражается в том, что образуются несколько сотен типов клеток специфического строения, отличающихся друг от друга. С биохимической точки зрения специализация клеток заключается в .синтезе определенных белков, свойственных только данному типу клеток. Биохимическая специализация клеток обеспечивается
С морфологической точки зрения дифференцирование выражается в том, что образуются несколько сотен типов клеток специфического строения, отличающихся друг от друга. С биохимической точки зрения специализация клеток заключается в .синтезе определенных белков, свойственных только данному типу клеток. Биохимическая специализация клеток обеспечивается 
У хордовых вскоре после гаструлянии небольшой участок спинной эктодермы в виде пластинки погружается в глубь зародыша, прогибается и образует нервную трубку с полостью внутри, заполненной жидкостью. Из клеток эктодермы развиваются кожные покровы с их производными (волосы, ногти, перья, копыта) и органы чувств. Из верхней части энтодермы образуется хорда, из нижней части — эпителий, выстилающий средние отделы кишечника, пищеварительные железы и органы дыхания. Из эктодермы, расположенной над хордой, развивается нервная трубка. Из мезодермы образуются мышцы, скелет, кровеносная система, половые железы, органы выделения и собственно кожа — дерма.
Эмбриональное развитие животных происходит или в материнском организме, или во внешней среде.
Гомология зародышевых листков подавляющего большинства животных — одно из доказательств единства животного мира.
Эмбриональная индукция. Эмбриональную индукцию можно определить как явление, при котором в процессе эмбриогенеза один зачаток влияет надругой, определяя путь его развития, и, кроме того, сам подвергается индуцирующему воздействию со стороны первого зачатка.
Зародышевое листки, их производные (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
| Название листка | Производные каждого листка |
| ЭКТОДЕРМА | Покровы тела (наружный эпителий, кожные железы, роговые чешуи, поверхностный слой зубов), нервная система, передний и задний отделы кишечника |
| ЭНТОДЕРМА | Эпителий средней кишки и пищеварительные железы, эпителий дыхательной системы |
| МЕЗОДЕРМА | Все мышечные, соединительные ткани, каналы выделительных органов, кровеносная система, часть тканей половых органов |
эмбриональное развитие (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
| стадия зиготы | дробление |
гаструляция | первичный гис то-и орга ногенез |
гисто- и органогенез |
| Однокле точный за родыш |
Формиро вание мно гоклеточ ного одно- слойного зароды ша — бла стулы |
Формиро вание двух- или трехслой ного заро дыша — гаструлы |
Формиро вание осе вых струк- -тур заро дыша |
Дифферен- цировка тканей, формиро вание ор ганов, рост заро дыша |
Постэмбриональный период развития
В момент рождения или выхода организма из яйцевых оболочек заканчивается эмбриональный и начинается постэмбриональный период развития. Постэмбриональное развитие может бытьпрямым sum непрямым и сопровождаться превращением {метаморфозом). При прямом развитии из .яйцевых оболочек или из тела матери выходит организм небольших размеров, но в нем заложены все основные органы, свойственные взрослому животному (пресмыкающиеся, птицы, млекопитающие). Постэмбриональное развитие у этих животных сводится в основном к росту и половому созреванию — дорепродуктивный период; размножению — репродуктивный период и старению — пострепрвдуктивный период.
Постэмбриональное развитие может бытьпрямым sum непрямым и сопровождаться превращением {метаморфозом). При прямом развитии из .яйцевых оболочек или из тела матери выходит организм небольших размеров, но в нем заложены все основные органы, свойственные взрослому животному (пресмыкающиеся, птицы, млекопитающие). Постэмбриональное развитие у этих животных сводится в основном к росту и половому созреванию — дорепродуктивный период; размножению — репродуктивный период и старению — пострепрвдуктивный период.
У организмов с малым содержанием желтка в яйце непрямое развитие сопровождается образованием личиночной стадии. Из яйца выходит личинка, обычно устроенная проще взрослого животного, со специальными личиночными органами, отсутствующими во взрослом состоянии. Личинка питается, растет, и, со временем личиночные органы заменяются органами, свойственными взрослым животным. При неполном метаморфозе замена личиночных органов происходит постепенно, без прекращения активного питания и перемещения организма (саранча, амфибии). Полный метаморфоз включает стадию куколки, в которой личинка преобразовывается во взрослое животное — имаго (бабочки).
Полный метаморфоз включает стадию куколки, в которой личинка преобразовывается во взрослое животное — имаго (бабочки).
Значение метаморфоза. 1) личинки могут самостоятельно питаться и растут, накапливая клеточный материал для формирования постоянных органов, свойственных взрослым животным; 2) свободноживущие личинки прикрепленных или паразитических, животных играют важную роль в расселении вида, в расширении ареала их обитания; 3) смена образа жизни или среды обитания в процессе индивидуального развития в результате того, что личиночные формы некоторых животных живут в иных условиях и имеют другие источники питания, чем взрослые особи, снижает интенсивность борьбы за существование внутри вида.
Эмбриональный период онтогенеза
☰
Онтогенез — это индивидуальное развитие организма. Он продолжается от момента зачатия, т. е. с образования зиготы, и до смерти. Эмбриональный период онтогенеза заканчивается с рождением детеныша или вылуплением его из яйца, икринки. После этого начинается постэмбриональное развитие организма.
После этого начинается постэмбриональное развитие организма.
У многоклеточных животных после своего образования зигота начинает многократно делиться путем митоза и образует бластулу. Данный процесс называется бластуляцией. Бластула — это шарообразная структура часто с полостью внутри, которую окружают мелкие клетки. Бластула по размеру не больше зиготы, т. е. клетки не росли, а при каждом последующем делении уменьшались в двое. Подобное деление называется дроблением.
Таким образом, первый этап эмбрионального развития — образование бластулы путем дробления зиготы.
Вторым этапом эмбрионального периода онтогенеза является образование гаструлы путем гаструляции — перемещении части клеток во внутрь полости бластулы с образованием двухслойного зародыша. При этом шарообразная структура становится похожа на чашеобразную. Бластула как бы впячивается с одной стороны во внутрь.
Внешний слой клеток гаструлы называется эктодермой, внутренний — энтодермой. Из этих клеток образуются различные ткани и органы. Также позже у большинства животных сформируется промежуточный слой клеток — мезодерма, который даст начало своим тканям и органам.
Из этих клеток образуются различные ткани и органы. Также позже у большинства животных сформируется промежуточный слой клеток — мезодерма, который даст начало своим тканям и органам.
После гаструляции наступает органогенез, когда формируются органы и их системы. У хордовых первым делом закладываются нервная трубка, хорда и кишечник.
Эмбрион на стадии закладки нервной трубки называется нейрулой.
Нервная система развивается из клеток эктодермы. Также из нее развиваются покровы тела и некоторые другие органы: эпидермис и его производные (когти, ногти, волосы, сальные и потовые железы), а также эпителий ротового и анального отверстий, слюнные железы, эмаль зубов.
Итак, из эктодермы формируются головной и спинной мозг, нервы, органы чувств и наружные покровы тела. Клетки мезодермы образуют скелет, мускулатуру, кровеносную, мочевыделительную, половую систему, формируют дентин зубов и дерму кожи. Из клеток энтодермы закладываются пищеварительная система (кроме слюнных желез), легкие и эпителий дыхательных путей.
Из клеток энтодермы закладываются пищеварительная система (кроме слюнных желез), легкие и эпителий дыхательных путей.
Таким образом, производные эктодермы выполняют в основном покровную и чувствительную функции, производные энтодермы – функции дыхания и питания, а производные мезодермы обеспечивают связи между частями зародыша – двигательную, опорную, транспортную и трофическую функции.
В процессе эмбриогенеза постепенно изменяется внешний вид зародыша. Он удлиняется, обособляются отделы тела. К концу эмбрионального периода онтогенеза плод становится похож на особь своего вида, хотя в начале развития все хордовые схожи между собой.
Конспект урока «Онтогенез. Индивидуальное развитие организма»
Урок № ……………
Цель: сформировать у учащихся представление об индивидуальном развитии организма.
Задачи. Обучающие – сформировать понятия онтогенез, эмбриогенез, бластула, гаструла, нейрула, гистогенез, органогенез, морула, бластоцель; познакомить с периодами онтогенеза; охарактеризовать процессы, протекающие на стадиях эмбриогенеза; познакомить с законом зародышевого сходства.
Развивающие – продолжить формирование навыков работы с книгой, с дополнительной научной литературой, немыми таблицами, интерактивной доской; развивать умение готовить сообщения, анализировать рисунки учебника, обобщать и делать выводы, выявлять закономерности.
Воспитывающие – аккуратное оформление схем в тетради, показать отрицательное влияние алкоголя, никотина, лекарственных препаратов, применяемых без назначения врача на эмбрион.
Оборудование. Презентация «Онтогенез. Эмбриональное развитие организма»
Ход урока.
I. Оргмомент.
II. Актуализация знаний.
Сегодня на уроке мы продолжаем изучение темы «Размножение и развитие организмов».
Фронтальный опрос.
Вспомните:
— Что называется размножением?
(Способность организма воспроизводить себе подобных).
— Какие Вы знаете способы размножения?
(Бесполое и половое).
— В чём различие между бесполым и половым размножением?
(При бесполом размножении принимает участие одна особь и дочерние организмы наследуют идентичную генетическую информацию материнской особи, а при половом –участие принимают два организма, которые производят гаметы и при слиянии гамет образуются особи с генетической информацией от обоих родителей ).
— Что такое гамета?
(Половая клетка ).
— Какие Вы знаете гаметы?
(Яйцеклетка и сперматозоид или спермии).
— Где они образуются?
(В половых железах).
— Что называется оплодотворением?
(Процесс слияния гамет).
— Что образуется в результате оплодотворения?
(Зигота).
— Что такое зигота?
(Это клетка с диплоидным набором хромосом, половина из которых получена от материнского организма , а половина – от отцовского).
Зигота – это одна клетка. Все живые организмы (грибы, растения, животные) начинают своё развитие с зиготы, то есть с одной клетки. О чём это свидетельствует?
Все живые организмы (грибы, растения, животные) начинают своё развитие с зиготы, то есть с одной клетки. О чём это свидетельствует?
(О родстве и единстве происхождения).
— Каким же образом из зиготы развивается целый организм? Из яйца курицы — цыплёнок, из икринки рыбы — малёк, из икринки лягушки – головастик, у млекопитающих – детёныш? Ведь зигота у большинства животных имеет микроскопические размеры, например, у млекопитающих 0,1 мм?
(Благодаря делению и росту клеток).
Правильно. Зигота претерпевает ряд изменений и благодаря вначале делению клеток, затем росту и дифференцировке клеток формируется организм. Деление клеток – рост клеток – дифференцировка клеток – это основа онтогенеза.
Цель сегодняшнего урока:
1. Познакомиться с понятием «онтогенез».
2. Познакомиться с типами и периодами онтогенеза.
3. Узнать, что такое эмбриональное развитие?
4. Выделить основные этапы эмбрионального развития.
Выделить основные этапы эмбрионального развития.
5. Установить какие изменения происходят на каждом этапе?
6. Установить влияние внешней среды на развитие зародыша.
План урока:
1. Онтогенез. Типы и этапы онтогенеза.
2. Эмбриональный период и его стадии.
— дробление
— бластула
— гаструла
— нейрула
— гистогенез и органогенез
3.Влияние частей развивающегося зародыша.
4. Влияние внешней среды на развитие зародыша.
III. Изучение нового материала.
ОНТОГЕНЕЗ.
Онтогенез ( греч.ontos – сущее, genesis — происхождение) – процесс, присущий любому живому организму, независимо от сложности его организации.
Онтогенезом, или индивидуальным развитием, называют весь период жизни с момента слияния половых клеток и образования зиготы до гибели организма.
Изучением вопросов, связанных с индивидуальным развитием организмов, занимается эмбриология (от греч. еmbryon – зародыш).
еmbryon – зародыш).
Исторические сведения:
Процесс появления и развития живых организмов интересовал людей с давних пор, но эмбриологические знания накапливались постепенно и медленно. Великий Аристотель, наблюдая за развитием цыпленка, предположил, что эмбрион образуется в результате смешения жидкостей, принадлежащих обоим родителям. Такое мнение продержалось в течение 200 лет. В XVII веке английский врач и биолог У. Гарвей проделал некоторые опыты для проверки теории Аристотеля. Будучи придворным врачом Карла I, Гарвей получил разрешение на использование для опытов оленей, обитающих в королевских угодьях. Гарвей исследовал 12 самок оленей, погибших в разные сроки после спаривания. Первый эмбрион, извлеченный из самки оленя через несколько недель после спаривания, был очень мал и совсем не похож на взрослое животное. У оленей, погибших в более поздние сроки, зародыши были крупнее, у них было большое сходство с маленькими, только что родившимися оленятами.
Так постепенно и накапливались знания по эмбриологии.
Основателем современной эмбриологии считается академик Российской Академии К.М.Бэр. В 1828 году он опубликовал сочинение «История развития животных», в котором доказывал, что человек развивается по единому плану со всеми позвоночными животными.
У простейших организмов тело которых состоит из одной клетки онтогенез совпадает с клеточным циклом, т.е. с момента появления, путем деления материнской клетки до следующего деления или смерти.
У многоклеточных видов размножающихся бесполым путем, онтогенез начинается с выделения группы клеток материнского организма (почкование гидры), которые делясь митозом, формируют новую особь со всеми ее системами и органами.
У тех видов, которые размножаются половым путем, онтогенез начинается с момента оплодотворения яйцеклетки и образования зиготы.
Онтогенез – это не просто рост маленькой особи до тех пор, пока она не превратится в большую. Это цепь строго определенных сложнейших процессов, в результате которых формируется присущие только особям данного вида особенности строения, жизненных процессов, способность к размножению. Заканчивается онтогенез процессами, закономерно ведущими к старению и смерти.
Заканчивается онтогенез процессами, закономерно ведущими к старению и смерти.
С генами родителей новая особь получает своего рода инструкцию о том, когда и какие, изменения должны происходить в организме, чтобы он смог успешно пройти весь жизненный путь.
ТИПЫ ОНТОГЕНЕЗА.
У животных выделяют три типа онтогенеза
Личиночный (встречается у насекомых, рыб, земноводных). Желтка в их яйцеклетках мало, и зигота быстро развивается в личинку, которая самостоятельно питается.
Яйцекладный (наблюдается у рептилий, птиц и яйцекладущих млекопитающих). Яйцеклетки данных видов живых организмов богаты желтком. Зародыш таких видов развивается внутри яйца.
Внутриутробный (у большинства млекопитающих и человека). При этом развивающийся зародыш задерживается в материнском организме, образуется временный орган – плацента, через который организм матери обеспечивает все потребности растущего эмбриона: дыхание, питание, выделение и т.
 д. Внутриутробное развитие заканчивается процессом деторождения.
д. Внутриутробное развитие заканчивается процессом деторождения.
ПЕРИОДЫ ОНТОГЕНЕЗА.
Любой тип онтогенеза у многоклеточных животных принять делить на 2 периода: эмбриональный и постэмбриональный.
Эмбриональный период начинается с оплодотворения и образования зиготы, заканчивается либо выходом личинки из оболочки, либо выходом особи из яйца, либо рождением особи.
Постэмбриональный период начинается с завершения эмбрионального периода. И включает в себя: половое созревание, взрослое состояние, старение и заканчивается смертью.
Сегодня мы остановимся поподробнее только на первом периоде онтогенеза – эмбриональном, или эмбриогенезе.
И рассмотрим мы развитие эмбриона на примере тех животных, чьи яйцеклетки содержат очень мало питательных веществ. К таким животным относятся плацентарные млекопитающие, в том числе и человек.
У вас на столах лежат листы с таблицей, которую вы будете заполнять по ходу моего объяснения.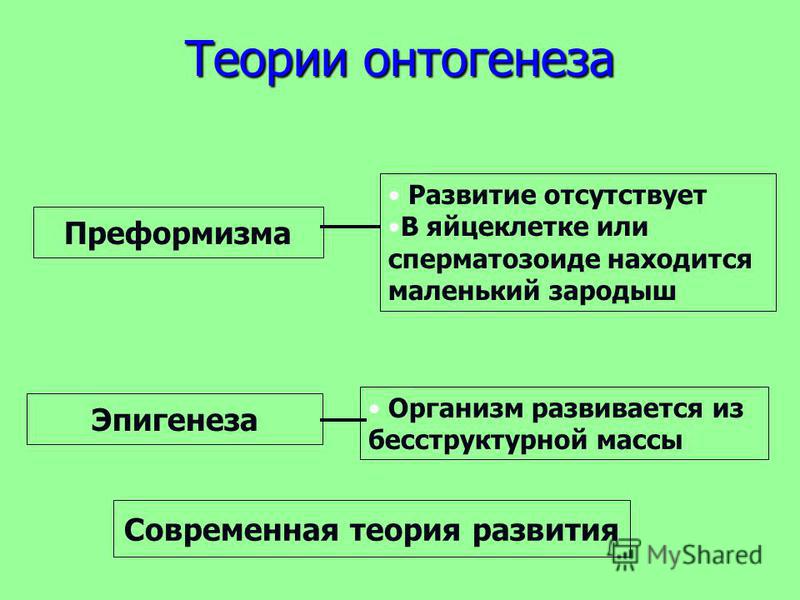
Основные этапы эмбрионального развития
Основные этапы
Особенности каждого этапа
Схематичный рисунок
1. Образование зиготы
Образуется при слиянии сперматозоида и яйцеклетки.
2. Образование бластулы
Дробление зиготы. Деление клеток, которые не сопровождается ростом. Образуется многоклеточный шар, состоящий из 32 клеток. Внутри шара находится полость — бластоцель
3. Образование гаструлы
Деление клеток на одном из полюсов бластулы и впячивание их внутрь бластоцели — гаструляция. Образование двух зародышевых листов – эктодермы и энтодермы, а затем развитие мезодермы.
4. Стадия нейрулы
Формирование важных частей зародыша — нервной трубки и хорды. Нервная трубка развивается из эктодермы, а хорда из мезодермы.
5. Закладка и формирование органов – органогенез
Процесс дифференцировки клеток и формирование органов.
Развитие организма начинается с одноклеточной стадии, которая происходит с момента слияния сперматозоида и яйцеклетки. Возникшее при оплодотворении ядро, обычно уже через несколько минут начинает делиться, вместе с ним делиться и цитоплазма. Образующиеся клетки, ещё сильно отличаются от клеток взрослого организма, называются бластомерами (от греч. blastos – зародыш,meros – часть). При делении бластомеров размеры их не увеличиваются, поэтому процесс деления носит название дробления. Дробление завершается образованием однослойного многоклеточного зародыша – бластулы. При дроблении клеток у всех животных – общий объем бластомеров на стадии бластулы не превышает объема зиготы. В итоге образуется многоклеточный шар, состоящий из 32 клеток. Внутри шара находится полость – бластоцель.
У человека на 6 день после оплодотворения бластула выходит из яйцевода в полость матки, а на седьмой день внедряется в ее стенку. Этот процесс называется имплантацией зародыша. После этого на одном из полюсов бластулы ее клетки начинают делиться быстрее, чем на другом, и впячиваться внутрь бластоцели. Это процесс получил название гаструляция. Вскоре из впячивания образуется второй внутренний слой клеток зародыша. Такой двухслойный шарик называется гаструлой. Наружная стенка гаструлы называется – наружным зародышевым листком или эктодермой, а внутренняя – внутренним зародышевым листком или энтодермой. Из клеток расположенных на границе между экто- и энтодермой , развивается третий зародышевый листок – мезодерма.
Этот процесс называется имплантацией зародыша. После этого на одном из полюсов бластулы ее клетки начинают делиться быстрее, чем на другом, и впячиваться внутрь бластоцели. Это процесс получил название гаструляция. Вскоре из впячивания образуется второй внутренний слой клеток зародыша. Такой двухслойный шарик называется гаструлой. Наружная стенка гаструлы называется – наружным зародышевым листком или эктодермой, а внутренняя – внутренним зародышевым листком или энтодермой. Из клеток расположенных на границе между экто- и энтодермой , развивается третий зародышевый листок – мезодерма.
Полость внутри гаструлы называется первичной кишкой, а отверстие, которое в нее ведет – первичным ртом.
Следующая за гаструлой стадия развития зародыша – нейрула. На этой стадии происходит формирование таких важных частей зародыша, как нервная трубка, и хорда. Нервная трубка развивается из эктодермы, а хорда из мезодермы.
Заключительным этапом является органогенез. Таким образом, уже на ранних стадиях эмбрионального периода онтогенеза из внешне одинаковых бластомеров развиваются различные по строению и функциям ткани, органы и системы. Этот процесс получил название дифференцировки клеток. Из каждого зародышевого листка формируются свои органы и системы, например,
Зародышевый лист
Органы
эктодерма
Нервная система, органы чувств, эпителий кожи, эмаль зубов
энтодерма
Мышечная ткань, соединительная ткань, кровеносная система, почки, половые железы
мезодерма
Эпителий средней кишки, пищеварительные железы — печень и поджелудочная железа, эпителий жабр и легких
ВЛИЯНИЕ УСЛОВИЙ ВНЕШНЕЙ СРЕДЫ НА РАЗВИТИЕ ЭМБРИОНА.
В эмбриональном периоде развитие любого организма зависит от условий окружающей среды. Причем в большей степени эта зависимость проявляется у плацентарных животных. Яйца птиц, например, практически изолированы от окружающей среды, а оптимальную температуру для зародыша обеспечивают родители при высиживании. Эмбриональное развитие млекопитающих находится в полной зависимости от материнского организма, т. к. от него получает все необходимое для жизни.
Яйца птиц, например, практически изолированы от окружающей среды, а оптимальную температуру для зародыша обеспечивают родители при высиживании. Эмбриональное развитие млекопитающих находится в полной зависимости от материнского организма, т. к. от него получает все необходимое для жизни.
Интенсивно делящиеся клетки зародыша весьма чувствительны к неблагоприятным воздействиям, которые могут привести к различным нарушениям в формирующимся организме. Опаснее всего воздействие химических веществ, способных проникать через плаценту в эмбрион. В частности к таким веществам относятся никотин, алкоголь, наркотики, действие радиоактивности, рентгеновых лучей, ядовитых веществ, различных медикаментов могут привести к очень тяжёлым последствиям — рождению ребёнка без рук, ног и даже без головы.
При нарушении почти любого звена эмбрионального развития возникают отклонения от нормального хода развития, т.е. аномалии.
Аномалии могут касаться органов кровообращения, дыхания, пищеварения, мочеполовой системы; возможно незаращение перегородок между предсердиями, образование добавочных селезёнок, удвоение почек и т. п
п
Наиболее опасными для развития зародыша являются первые три месяца беременности. В этот период плод особенно чувствителен к вирусным инфекциям, поскольку плаценты еще нет. Она формируется к концу третьего месяца беременности. Например, такое почти безопасное для взрослых и детей заболевание, как краснуха, может привести к рождению ребенка с пороком сердца, глухотой, умственной отсталостью, если мать заболеет этой болезнью в начале беременности.
Вредное влияние на развитие эмбриона оказывает употребление его родителями алкоголя, наркотиков, курение табака. Алкоголь и никотин угнетают клеточное дыхание. Недостаточное снабжение кислородом приводит к тому, что в формирующихся органах образуется меньшее количество клеток, органы оказываются недоразвитыми. Особенно чувствительна к недостатку кислорода нервная ткань. Употребление будущей матерью алкоголя, наркотиков, курение табака, злоупотребление лекарствами часто приводят к необратимому повреждению эмбриона и последующему рождению детей с умственной отсталостью или врожденными уродствами. В легких случаях наблюдается расторможенность ребенка: несобранность, неспособность к систематической работе, нерациональная двигательная активность, низкий уровень произвольного внимания, а также очень плохая память.
В легких случаях наблюдается расторможенность ребенка: несобранность, неспособность к систематической работе, нерациональная двигательная активность, низкий уровень произвольного внимания, а также очень плохая память.
Эмбрион очень чувствителен к лекарственным препаратам. Поэтому беременные женщины должны их применять строго по назначению врача.
Не меньшую опасность для развития зародыша представляет загрязнение среды обитания различными химическими веществами или облучение ионизирующей радиацией.
Составление схемы факторов влияющих на плод в тетради.
— А сейчас, я вам прочту отрывок из стихотворения Али Ибн Сина «Поэма о медицине»
О детях, находящихся ещё в утробе матери:
Как следует, о том веду я речь.
Дитя в утробе матери беречь.
Ничто зловредное его пусть не коснётся,
Пусть мать питается не как придётся,
А ест еду и влагу с пользой пьёт,
Так, чтоб нормально развивался плод.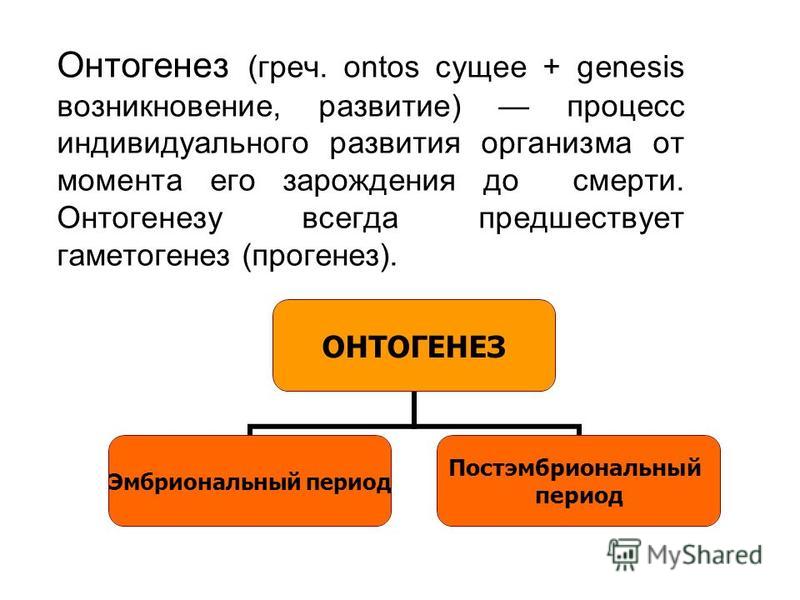
Пускай отбросов в пище будет мало,
Чтоб кровь она при этом очищала.
— Как вы понимаете строки этого стихотворения?
Обсуждение стихотворения.
Учитель: Как называется процесс развития зародыша в матке женщины?
Ученик: Этот процесс называется беременностью.
Учитель: Вы становитесь взрослыми. И вам всем уже известно, что любая половая близость, может завершиться беременностью. Девочки, девушки, и даже взрослые женщины стоят перед выбором сохранить беременность или её прервать. Прежде чем пойти на страшный шаг убийства, а это действительно убийство, подумайте! А может это ваша последняя беременность и после сделанного аборта, вы никогда не будите иметь детей, и не познаете счастья материнства. Возможно, этот ребёнок был бы вашей опорой в жизни, был бы самым умным, талантливым. Действительно, для любого нормального человека уничтожение живой жизни безнравственно, греховно. Необходимо отметить, что ответственность за аборт ложиться на двоих – мужчину и женщину.
Необходимо отметить, что ответственность за аборт ложиться на двоих – мужчину и женщину.
— Сейчас вы прослушаете «Дневник нерожденного ребенка».
— А я призываю вас только к одному: задумайтесь!
IV. Закрепление.
Подведём итоги изученного на уроке.
Фронтальный опрос или кроссворд.
— Что такое онтогенез? — Какие типы онтогенеза вы знаете?
— Какие периоды он включает?
— Эмбриогенез – это……?
— Назовите стадии эмбриогенеза?
— Кратко охарактеризуйте каждую из них? ( использование таблицы)
— Перечислите, какие вам известны зародышевые листки и что в последствии из них образуется?
V. Домашнее задание.
§35, 36 «Биология»А.А.Каменский.
Сообщение : «Почему же мы стареем…?»
ОСНОВНЫЕ ЭТАПЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Основные этапы
Особенности каждого этапа
Схематичный рисунок
1. Образование зиготы
Образование зиготы
2. Образование бластулы
3. Образование гаструлы
4. Стадия нейрулы
5. Закладка и формирование органов – гистогенез
Кроссворд «Онтогенез. Эмбриональное развитие»
9
10
7
8
5
3
4
2
6
1
1. Клетки, образующиеся при делении зиготы и отличающиеся от клеток взрослого организма.
Клетки, образующиеся при делении зиготы и отличающиеся от клеток взрослого организма.
2. Основатель современной эмбриологии.
3. Первая клетка нового организма.
4. Этап, в процессе которого происходит закладка и формирование органов.
5. Новый организм, который развивается из оплодотворенной яйцеклетки.
6. Тип онтогенеза, характерный для насекомых, рыб и земноводных.
7. Этап, в результате которого происходит формирование важных частей зародыша.
8. Полость, находящаяся внутри шара образованного бластомерами.
9. Период онтогенеза, который начинается с оплодотворения и заканчивается появлением новой особи.
10. Временный орган в материнском организме, обеспечивающий эмбриону дыхание, питание, выделение и т.д.
Кроссворд «Онтогенез. Эмбриональное развитие»
9
э
м
б
р
и
о
10
п
л
а
ц
е
н
т
а
7
а
н
8
б
л
а
с
т
о
ц
е
л
ь
ь
й
н
5
р
ы
з
у
й
а
л
3
4
о
р
г
а
н
о
г
е
н
е
з
о
и
д
2
г
6
л
и
ч
и
н
о
ч
н
ы
й
1
б
л
а
с
т
о
м
е
р
ш
э
т
р
а
1. Клетки, образующиеся при делении зиготы и отличающиеся от клеток взрослого организма.
Клетки, образующиеся при делении зиготы и отличающиеся от клеток взрослого организма.
2. Основатель современной эмбриологии.
3. Первая клетка нового организма.
4. Этап, в процессе которого происходит закладка и формирование органов.
5. Новый организм, который развивается из оплодотворенной яйцеклетки.
6. Тип онтогенеза, характерный для насекомых, рыб и земноводных.
7. Этап, в результате которого происходит формирование важных частей зародыша.
8. Полость, находящаяся внутри шара образованного бластомерами.
9. Период онтогенеза, который начинается с оплодотворения и заканчивается появлением новой особи.
10. Временный орган в материнском организме, обеспечивающий эмбриону дыхание, питание, выделение и т.д.
Начало формы
Гены, работающие на самых консервативных стадиях онтогенеза, оказались самыми многофункциональными
Модель «песочных часов» предполагает, что средние стадии эмбрионального развития, когда закладывается характерный для данного типа животных план строения, обладают повышенной эволюционной консервативностью по сравнению с ранними и поздними стадиями. Сравнение экспрессии генов на разных этапах развития у восьми видов, представляющих разные эволюционные ветви хордовых, подтвердило приложимость модели «песочных часов» к подтипу позвоночных и, в несколько меньшей степени, ко всему типу хордовых. Кроме того, выяснилось, что гены, работающие на средних стадиях развития, отличаются повышенной многофункциональностью: многие из них выполняют разнообразные функции на разных стадиях развития и в разных частях организма. Это может быть одной из причин консервативности средних стадий эмбрионального развития.
Сравнение экспрессии генов на разных этапах развития у восьми видов, представляющих разные эволюционные ветви хордовых, подтвердило приложимость модели «песочных часов» к подтипу позвоночных и, в несколько меньшей степени, ко всему типу хордовых. Кроме того, выяснилось, что гены, работающие на средних стадиях развития, отличаются повышенной многофункциональностью: многие из них выполняют разнообразные функции на разных стадиях развития и в разных частях организма. Это может быть одной из причин консервативности средних стадий эмбрионального развития.
Давно замечено, что эмбрионы хордовых животных сильнее всего похожи друг на друга на средних стадиях эмбрионального развития, особенно на стадии фарингулы (см. Pharyngula). В это время формируются характерные для типа хордовых морфологические структуры (сомиты, спинная нервная трубка, хорда, жаберные щели), в совокупности формирующие то, что называют «планом строения» хордовых. Данный этап развития, получивший название «филотипической стадии», обладает повышенной эволюционной консервативностью (то есть мало меняется в ходе эволюции) по сравнению с более ранними и более поздними этапами эмбриогенеза. Эти наблюдения отражены в модели «песочных часов развития» (рис. 1).
Эти наблюдения отражены в модели «песочных часов развития» (рис. 1).
Молекулярные исследования показали, что консервативность филотипической стадии прослеживается не только на морфологическом уровне, но и на уровне экспрессии генов. Профили экспрессии у эмбрионов разных видов, относящихся к одному типу, оказались наиболее похожими друг на друга именно на филотипической стадии. Правда, до сих пор такие исследования проводились лишь на очень ограниченном числе видов (например, на четырех, см.: N. Irie, S. Kuratani, 2011. Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis), и поэтому полной уверенности в том, что модель «песочных часов» приложима ко всему типу хордовых, не было.
Похожая закономерность была замечена и в онтогенезе других типов животных (нематод, членистоногих, кольчатых червей), у которых тоже, по-видимому, в развитии есть консервативная «филотипическая» стадия, когда закладывается план строения, характерный для данного типа (подробнее см. в новости Смысл консервативной стадии зародышевого развития начинает проясняться, «Элементы», 31.03.2017).
в новости Смысл консервативной стадии зародышевого развития начинает проясняться, «Элементы», 31.03.2017).
Идея о существовании универсальной филотипической стадии, общей для разных типов животных, не подтверждается молекулярными данными: по-видимому, у каждого типа она индивидуальна, как и план строения. Впрочем, тут важно иметь в виду, что в самих определениях понятий «тип» и «план строения» используется кольцевая логика: тип определяют как монофилетическую группу организмов, обладающих данным планом строения, а планом строения называют совокупность морфологических особенностей, характеризующую данный тип. Но, тем не менее, объективный факт состоит в том, что в развитии многих организмов (причем не только животных, но и растений и даже грибов; см.: М. Гельфанд. Песочные часы в биологии развития) средние стадии развития обладают пониженной изменчивостью по сравнению с ранними и поздними.
Причины этой закономерности точно не известны, хотя есть правдоподобные и аргументированные гипотезы (см. , например, рис. 1, процитированную выше новость и статью N. Irie, S. Kuratani, 2014. The developmental hourglass model: a predictor of the basic body plan?).
, например, рис. 1, процитированную выше новость и статью N. Irie, S. Kuratani, 2014. The developmental hourglass model: a predictor of the basic body plan?).
В новом масштабном исследовании, результаты которого опубликованы в журнале Nature Ecology & Evolution, большой коллектив китайских и японских исследователей сделал важный шаг к разгадке тайны «песочных часов». Авторы измерили экспрессию всех генов на разных стадиях развития у восьми видов с прочтенными геномами. Эти виды представляют все три подтипа хордовых (головохордовые, оболочники и позвоночные), а также несколько важнейших эволюционных ветвей позвоночных (костные рыбы, амфибии, млекопитающие, птицы и черепахи). Для полноты картины не хватает, пожалуй, хрящевых рыб и круглоротых, но и без того в данном исследовании разнообразие хордовых охвачено намного полнее, чем в прежних работах (рис. 2).
Полученные профили экспрессии затем сравнивали между собой, чтобы количественно оценить степень консервативности разных стадий развития. Как правильно проводить такие сравнения — вопрос крайне непростой, и среди ученых на сегодняшний день нет единой точки зрения на этот счет. Поэтому авторы на всякий случай использовали несколько разных подходов. Например, можно сравнивать экспрессию только тех генов, которые есть у каждого вида в единственном экземпляре. Тогда для каждого гена, например, лягушки можно будет найти один и только один соответствующий ген в геноме мыши, асцидии, ланцетника и т. д. Но таких генов не очень много (всего 1704, что составляет менее 10% от общего числа генов в геноме среднестатистического хордового). Другой вариант — включить в анализ также и гены, которые есть у большинства видов, но не обязательно в единственном экземпляре (у кого-то данный ген утрачен, у кого-то он присутствует в виде нескольких различающихся копий — паралогов). В этом случае можно, например, считать, что у вида, вообще не имеющего данного гена, уровень его экспрессии на всех стадиях равен нулю, а у вида, имеющего несколько паралогов, брать для каждой стадии либо средний, либо суммарный уровень экспрессии всех паралогов.
Как правильно проводить такие сравнения — вопрос крайне непростой, и среди ученых на сегодняшний день нет единой точки зрения на этот счет. Поэтому авторы на всякий случай использовали несколько разных подходов. Например, можно сравнивать экспрессию только тех генов, которые есть у каждого вида в единственном экземпляре. Тогда для каждого гена, например, лягушки можно будет найти один и только один соответствующий ген в геноме мыши, асцидии, ланцетника и т. д. Но таких генов не очень много (всего 1704, что составляет менее 10% от общего числа генов в геноме среднестатистического хордового). Другой вариант — включить в анализ также и гены, которые есть у большинства видов, но не обязательно в единственном экземпляре (у кого-то данный ген утрачен, у кого-то он присутствует в виде нескольких различающихся копий — паралогов). В этом случае можно, например, считать, что у вида, вообще не имеющего данного гена, уровень его экспрессии на всех стадиях равен нулю, а у вида, имеющего несколько паралогов, брать для каждой стадии либо средний, либо суммарный уровень экспрессии всех паралогов. Авторы перепробовали все эти способы. Результаты, полученные разными методами, в общих чертах получились сходными, хотя и различающимися в деталях.
Авторы перепробовали все эти способы. Результаты, полученные разными методами, в общих чертах получились сходными, хотя и различающимися в деталях.
Для каждого вида и для каждой стадии развития была оценена степень сходства по профилю генной экспрессии с разными стадиями других видов. Самыми консервативными считались те стадии, для которых было выявлено наибольшее сходство с другими видами (неважно, с какими именно стадиями их развития — ведь строгое соответствие между разными стадиями развития, например, асцидии и мыши провести затруднительно).
В итоге для каждого из восьми видов были несколькими разными способами построены графики, подобные изображенным на рис. 3.
Полученные результаты подтвердили, что средние стадии развития у позвоночных (причем не только стадия фарингулы, но и ряд предшествующих и последующих этапов) являются более консервативными, чем начальные и завершающие этапы эмбриогенеза. Таким образом, модель «песочных часов» подтвердилась для подтипа позвоночных.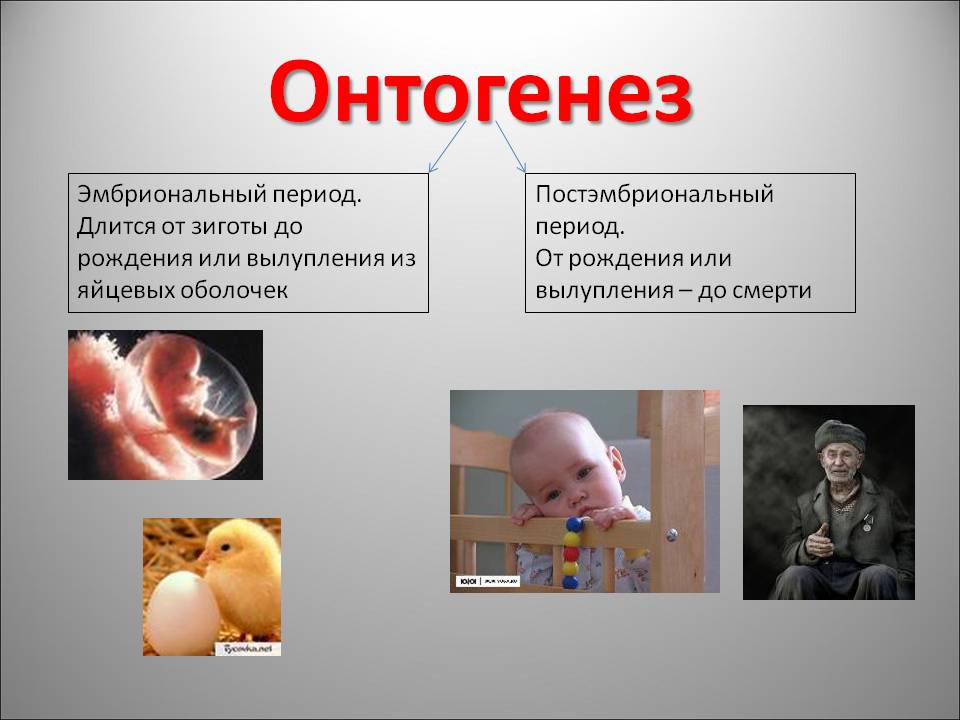
Что касается всего типа хордовых в целом, то для него эта модель в принципе тоже подтвердилась, но не так уверенно. Картинку портит асцидия, у которой вообще не очень понятно, что считать филотипической стадией: в ее развитии нет такого этапа, на котором одновременно присутствуют все четыре главных элемента «плана строения хордовых» (сегментированная мускулатура, нервная трубка, хорда, жаберные щели). Развитие оболочников претерпело вторичную модификацию и, скорее всего, сильно отличается от развития общего предка хордовых (см.: Причина особенностей генома оболочников — детерминированность их эмбрионального развития, «Элементы», 01.06.2014). Что, кстати, говорит о том, что эволюционная консервативность средних (и любых других) этапов развития далеко не абсолютна.
Но всё же в пределах подтипа позвоночных средние этапы эмбриогенеза сохраняют удивительное постоянство на протяжении сотен миллионов лет — и это несмотря на самые радикальные изменения образа жизни и строения как взрослого организма, так и начальных стадий развития.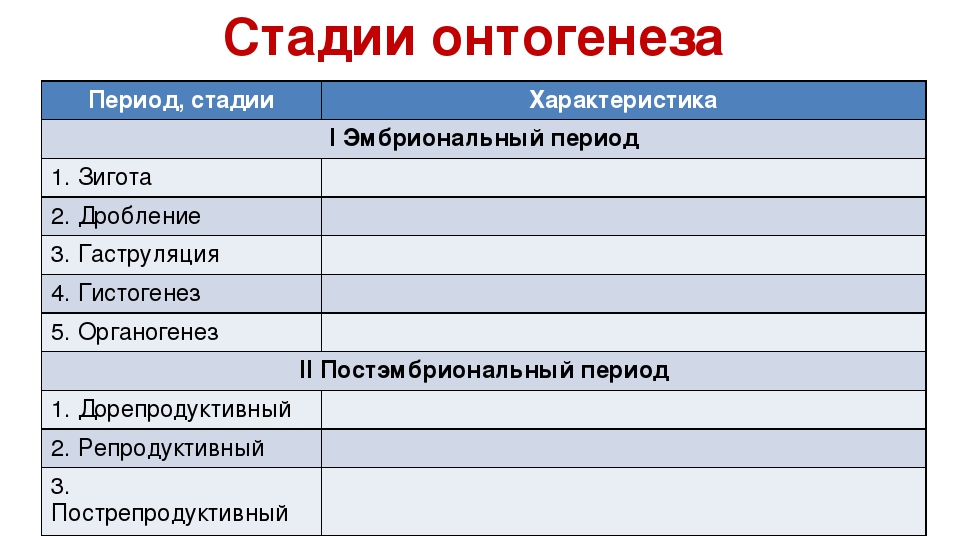
Чтобы разгадать причину этого постоянства, авторы решили выяснить, чем отличаются гены, работающие на средних стадиях эмбриогенеза, от генов, работающих на ранних и поздних стадиях.
Для каждой стадии развития (и для каждого вида) было подсчитано количество генов, экспрессия которых достоверно повышена или понижена именно на этой стадии. Оказалось, что число таких генов минимально как раз на средних, самых консервативных стадиях развития (рис. 4).
Выяснилось также, что среди генов, работающих на средних стадиях развития, повышена доля генов, которые экспрессируются, во-первых, во многих разных тканях и органах эмбриона, во-вторых — на многих разных этапах развития. Среди них также повышена доля генов, о которых известно, что они необходимы для нормального развития зародыша. Кроме того, для белков, кодируемых этими генами, характерно в среднем большее число белок-белковых взаимодействий (это значит, что они, скорее всего, вовлечены в большее число различных сигнальных и регуляторных каскадов).
Всё это, по-видимому, говорит о том, что для генов, контролирующих средние стадии развития, характерна повышенная плейотропия (многофункциональность). Многие из них задействованы не только в бурных морфогенетических процессах филотипической стадии развития, но и в других процессах на других стадиях. По-видимому, эти гены чаще других привлекались в ходе эволюции для выполнения каких-то новых функций (примеры того, как на старые регуляторные гены могут «навешиваться» новые функции, рассмотрены в новостях Ген, работающий в мышцах и костях, у обезьян стал регулировать развитие мозга, «Элементы», 14.11.2016; Вирус, встроившийся в геном наших предков, влияет на работу мозга, «Элементы», 19.11.2013; Найден ген, отвечающий за эволюцию окраски у бабочек, «Элементы», 31.08.2011).
Авторы предполагают, что повышенная плейотропия генов, работающих на филотипической стадии, является одной из причин консервативности этого этапа развития (рис. 5).
Множественность функций должна ограничивать эволюционную свободу гена: многофункциональный ген трудно изменить так, чтобы «подкрутить» только одну из его функций, не повредив остальным. Впрочем, до сих пор считалось, что данное рассуждение справедливо в первую очередь для белок-кодирующих участков. Хорошо известно, что многофункциональность гена действительно повышает консервативность его белок-кодирующей части. В данном случае, однако, речь идет уже о следующем, более высоком уровне консервативности — о затрудненности эволюционных изменений уровня экспрессии, определяемого регуляторными участками гена. Влияние многофункциональности на консервативность регуляторных участков — идея довольно-таки новая и нуждающаяся в дополнительных подтверждениях и объяснениях.
Впрочем, до сих пор считалось, что данное рассуждение справедливо в первую очередь для белок-кодирующих участков. Хорошо известно, что многофункциональность гена действительно повышает консервативность его белок-кодирующей части. В данном случае, однако, речь идет уже о следующем, более высоком уровне консервативности — о затрудненности эволюционных изменений уровня экспрессии, определяемого регуляторными участками гена. Влияние многофункциональности на консервативность регуляторных участков — идея довольно-таки новая и нуждающаяся в дополнительных подтверждениях и объяснениях.
Можно также предположить, что многофункциональные гены, работающие на филотипической стадии, настолько важны для нормального развития организма, что система регуляции их экспрессии приобрела в ходе эволюции повышенную помехоустойчивость (например, благодаря дополнительным регуляторным контурам, дублирующим и подстраховывающим друг друга; см. Избыточные регуляторы делают развитие эмбриона помехоустойчивым, «Элементы», 03. 06.2010). В таком случае, возможно, их замедленная эволюция объясняется зарегулированностью и пониженным уровнем изменчивости, за которую мог бы зацепиться естественный отбор.
06.2010). В таком случае, возможно, их замедленная эволюция объясняется зарегулированностью и пониженным уровнем изменчивости, за которую мог бы зацепиться естественный отбор.
Главный вопрос, оставшийся без ответа, состоит, конечно же, в том, почему самыми многофункциональными оказались именно гены, работающие на филотипической стадии. Можно предположить, что гены, умеющие контролировать закладку разнообразных сложных органов, просто в силу своих «профессиональных навыков» особенно удобны для того, чтобы навешивать на них новые функции — например, регуляцию развития каких-то новых структур, возникающих в ходе адаптации к меняющейся среде. Но пока это лишь общие рассуждения, нуждающиеся в экспериментальной проверке и конкретизации.
Источник: Haiyang Hu, Masahiro Uesaka, Song Guo, Kotaro Shimai, Tsai-Ming Lu, Fang Li, Satoko Fujimoto, Masato Ishikawa, Shiping Liu, Yohei Sasagawa, Guojie Zhang, Shigeru Kuratani, Jr-Kai Yu, Takehiro G. Kusakabe, Philipp Khaitovich, Naoki Irie and the EXPANDE Consortium. Constrained vertebrate evolution by pleiotropic genes // Nature Ecology & Evolution. Published online 25 September 2017.
Constrained vertebrate evolution by pleiotropic genes // Nature Ecology & Evolution. Published online 25 September 2017.
См. также:
Смысл консервативной стадии зародышевого развития начинает проясняться, «Элементы», 31.03.2017.
Александр Марков
Индивидуальное развитие организмов
Вопросы внутри параграфа: Какие этапы проходит организмов в своем развитии?
1)Эмбриональный
2)Постэмбриональный:
-Репродуктивный
-Пострепродуктивный
-Старость
-Смерть
Почему гаметофит считают половым, а спорофит – бесполым поколением? Гаметофит – это гаплоидная многоклеточная стадия в жизненном цикле растений и водорослей, развивающаяся из спор и производящая половые клетки (гаметы), а споры образуются в результате репродуктивного деления (мейоза) и всегда гаплоидны, то есть содержат одинарный гаплоидный набор хромосом. Спорофит – это диплоидное поколение высших растений, образующееся при объединении генетических материалов женской и мужской гамет.
Спорофит – это диплоидное поколение высших растений, образующееся при объединении генетических материалов женской и мужской гамет.
Стр. 97. Вопросы и задания после §
1. Что называют онтогенезом?
Онтогенез – это индивидуальное развитие организмов от момента образования зиготы до смерти организма.
2. Почему считают, что онтогенез – запрограммированный процесс?
С открытием в 20 веке генов и хромосом на смену примитивным представлениям о развитии, пришло учение о генетической информации, направляющей ход онтогенеза. Все признаки организма определяются последовательностью нуклеотидов в ДНК клетки. В ДНК «записана» программа индивидуального развития особи.
3. Какие этапы онтогенеза животных встречаются в природе? Приведите примеры организмов с различными типами онтогенеза.
Эмбриональный период – отрезок времени от начала дробления зиготы и до выхода организма из яйцевых или зародышевых оболочек. Постэмбриональный этап начинается после рождения либо выхода из яйцевых оболочек. У организмов выделают два типа онтогенеза: прямой и непрямой. Непрямой тип характеризуется тем, что яйцеклетка содержит мало питательных веществ, необходимых для завершения развития. Поэтому имеется стадия личинки, на которой формируются особые личиночные приспособления. Например, для головастика характерна обтекаемая форма тела, жабры, хвост, боковая линия, и другие черты для обитания в воде. Это личиночный тип онтогенеза. Прямой тип онтогенеза характерен для птиц, пресмыкающихся, млекопитающих. Стадии личинки отсутствует. У птиц и пресмыкающихся яйцеклетки богаты желтком, зародыш развивается в плотных яйцевых оболочках, он хорошо защищен и снабжен питательными веществами. У млекопитающих развитие идет в утробе матери, такое развитие называется внутриутробным.
Постэмбриональный этап начинается после рождения либо выхода из яйцевых оболочек. У организмов выделают два типа онтогенеза: прямой и непрямой. Непрямой тип характеризуется тем, что яйцеклетка содержит мало питательных веществ, необходимых для завершения развития. Поэтому имеется стадия личинки, на которой формируются особые личиночные приспособления. Например, для головастика характерна обтекаемая форма тела, жабры, хвост, боковая линия, и другие черты для обитания в воде. Это личиночный тип онтогенеза. Прямой тип онтогенеза характерен для птиц, пресмыкающихся, млекопитающих. Стадии личинки отсутствует. У птиц и пресмыкающихся яйцеклетки богаты желтком, зародыш развивается в плотных яйцевых оболочках, он хорошо защищен и снабжен питательными веществами. У млекопитающих развитие идет в утробе матери, такое развитие называется внутриутробным.
4. Чем характеризуется эмбриональный период онтогенеза животных?
Эмбриональный период – отрезок времени от начала дробления зиготы и до выхода организма из яйцевых или зародышевых оболочек.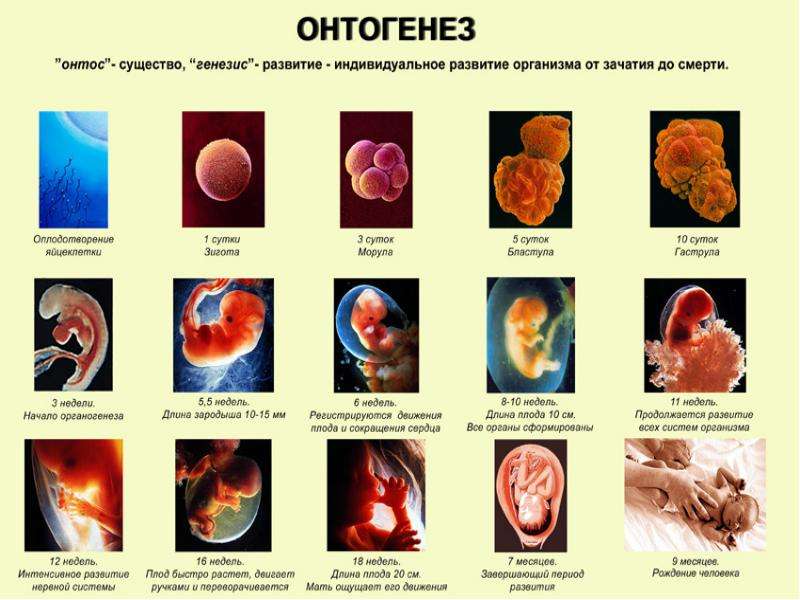 Спустя некоторое время после оплодотворения зигота начинает дробиться. Благодаря следующим друг за другом митотическим делениям из одной крупной оплодотворенной яйцеклетки образуется множество более мелких клеток – бластомеров. В центральной части зародыша образуется полость и наступает стадия бластулы.
Спустя некоторое время после оплодотворения зигота начинает дробиться. Благодаря следующим друг за другом митотическим делениям из одной крупной оплодотворенной яйцеклетки образуется множество более мелких клеток – бластомеров. В центральной части зародыша образуется полость и наступает стадия бластулы.
Стадии эмбрионального периода:
-Бластуляция – однойслойный зароды
-Гаструляция – двуслойный зародыш (энтодерама и эктодерама)
-Нейруляция – трехслойный зародыш (энтодерма, эктодерма, мезодерма)
-Органогенез – образование органов (первичные: хорда, кишечная трубка, нервная трубка)
Постепенно закладываются ткани, формируются органы и системы органов. У семенных растений эмбриональным считается период от оплодотворения яйцеклетки до прорастания зародыша.
5. Что означает термин «чередование поколений» у растений?
Чередование поколений – это смена поколений в жизненном цикле растения, половое гаплоидное поколение – гаметофит, сменяет бесполое диплоидное поколение – спорофит.
6. В чем особенность постэмбрионального этапа у растений?
У растений, в отличие от большинство животных, формирование органов и тканей осуществляется на только во время развития зародыша, но и на протяжении всего постэмбрионального периода.
Онтогенез и филогенез
Процесс индивидуального развития особи от начала её существования и до конца жизни называют онтогенезом.
У бактерий и простейших онтогенез практически совпадает с клеточным циклом.
У многоклеточных видов, которые размножаются бесполым путём, онтогенез начинается с момента выделения группы клеток материнского организма, которые затем делятся митозом и формируют новую особь со всеми системами и органами.
У
тех видов, которые размножаются половым путём, онтогенез начинается с момента
оплодотворения яйцеклетки и образования зиготы — первой клетки нового
организма.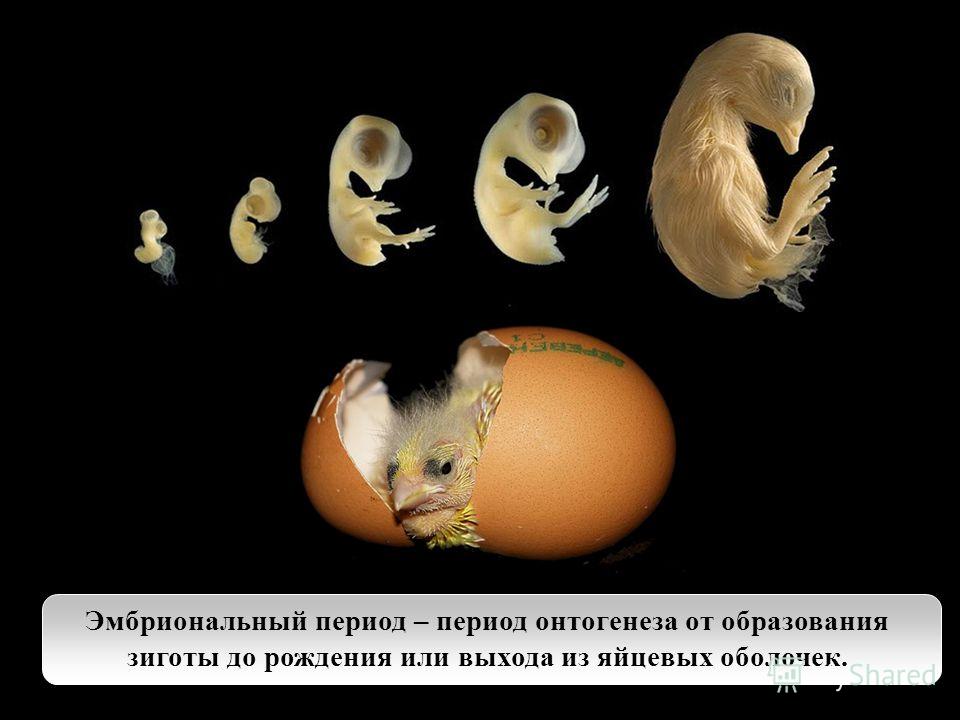
У животных выделяют три типа онтогенеза: личиночный, яйцекладный и внутриутробного развития.
Личиночный тип развития.
Он встречается, например, у насекомых, рыб, земноводных. Желтка в яйцеклетках этих организмов мало, и зигота быстро развивается в личинку, которая самостоятельно питается и растёт. Затем, по прошествии какого-то времени, происходит метаморфоз, — то есть превращение личинки во взрослую особь.
У некоторых видов наблюдается даже целая цепочка превращений из одной личинки в другую и только потом — во взрослую особь.
Например, у кузнечика личинка питается, растёт, несколько раз линяет. После окончательной линьки она становится взрослой половозрелой особью с крыльями и больше не растёт.
Яйцекладный тип онтогенеза
Он
наблюдается у рептилий, птиц и яйцекладущих млекопитающих, яйцеклетки которых
богаты желтком. Зародыш таких видов развивается внутри яйца; личиночная стадия
отсутствует.
Питание зародыша обеспечивает желток. Содержимое яйца окружено двумя подскорлупковыми оболочками, внутренней и наружной. Снаружи находится скорлупа. После откладки яйца на его тупом конце постепенно образуется воздушная камера.
Однако у большинства млекопитающих яйцо не образуется, а зародыш развивается внутри тела самки в половом органе — матке.
Внутриутробный тип онтогенеза
Он наблюдается у большинства млекопитающих, в том числе и у человека. При этом развивающийся зародыш задерживается в материнском организме, образуется временный орган — плацента, через который организм матери обеспечивает все потребности растущего эмбриона: дыхание, питание, выделение и др. Внутриутробное развитие оканчивается процессом деторождения.
Периоды онтогенеза
Любой
вид индивидуального развития у многоклеточных животных принято делить на два
периода: эмбриональный и постэмбриональный.
Эмбриональный период начинается с оплодотворения и представляет собой процесс формирования сложного многоклеточного организма, в котором представлены все системы органов.
Заканчивается этот период выходом личинки из своих оболочек (при личиночном типе онтогенеза), выходом особи из яйца (при яйцекладном типе) или рождением особи (при внутриутробном типе).
Постэмбриональный период начинается с завершения эмбрионального. Он включает в себя половое созревание, взрослое состояние, старение, которое заканчивается смертью.
Этапы эмбрионального развития на примере ланцетника.
Вспомним, что ланцетник ― это примитивное морское животное из семейства ланцетниковых. Ланцетник рассматривается как промежуточное звено между позвоночными и беспозвоночными животными.
В
результате оплодотворения гаплоидные ядра половых клеток сперматозоида и
яйцеклетки сливаются. Так возникает одноклеточная стадия развития организма — зигота,
которая содержит диплоидный набор хромосом.
Так возникает одноклеточная стадия развития организма — зигота,
которая содержит диплоидный набор хромосом.
Зигота является началом нового организма. Она делится митозом.
Митотические деления называются делениями дробления.
Дробление (бластуляция) ― это процесс образования многоклеточного зародыша.
Первое деление происходит в вертикальной плоскости, и клетка делится на две одинаковые клетки с диплоидным набором хромосом. Их называют бластомерами.
Бластомеры не расходятся, а делятся ещё раз, и образуется уже 4 клетки.
Третье деление происходит в горизонтальной плоскости, и из четырёх образуется 8 бластомеров.
Далее продольные и поперечные деления сменяют друг друга. Возникает все больше бластомеров, которые не успевают расти.
После
нескольких делений, когда число бластомеров достигает 32, они образуют полый
шарик со стенками в один ряд клеток. Этот шарик получил название бластулы.
Этот шарик получил название бластулы.
Бластула имеет внутри полость (пустое пространство). Эту полость называют первичной полостью тела или бластоцелью.
У человека на шестой день после оплодотворения бластула выходит из яйцевода в полость матки, а затем внедряется в её стенку. Этот процесс называется имплантацией зародыша.
После этого на одном из полюсов бластулы её клетки начинают делится быстрее, чем на другом, и впячиваться внутрь бластоцели. Этот процесс получил название гаструляции.
Так формируется второй внутренний слой клеток ― зародышевый листок. Такой двуслойный шарик называется гаструлой.
Гаструла состоит из двух зародышевых листков. Верхние клетки составляют наружный зародышевый листок ― эктодерму.
А
внутренние ― внутренний зародышевый листок ― энтодерму.
Полость, которая образовалась внутри гаструлы, является первичной кишкой.
А отверстие, ведущее в первичную кишку, — первичным ртом.
У позвоночных животных на месте первичного рта в процессе эмбриогенеза образуется анальное отверстие, а настоящий (вторичный рот) возникает на противоположном полюсе зародыша. Поэтому млекопитающих (как и всех хордовых животных) относят к вторичноротым.
Между энтодермой и эктодермой образуется третий (средний) зародышевый листок — мезодерма.
Следующая стадия развития зародыша называется нейрулой.
На этой стадии происходит формирование нервной трубки и хорды.
У большинства хордовых хорда присутствует только в период эмбрионального развития, в дальнейшем замещаясь позвоночником.
У
низших хордовых она сохраняется всю жизнь, выполняя опорную функцию, например,
как у ланцетника.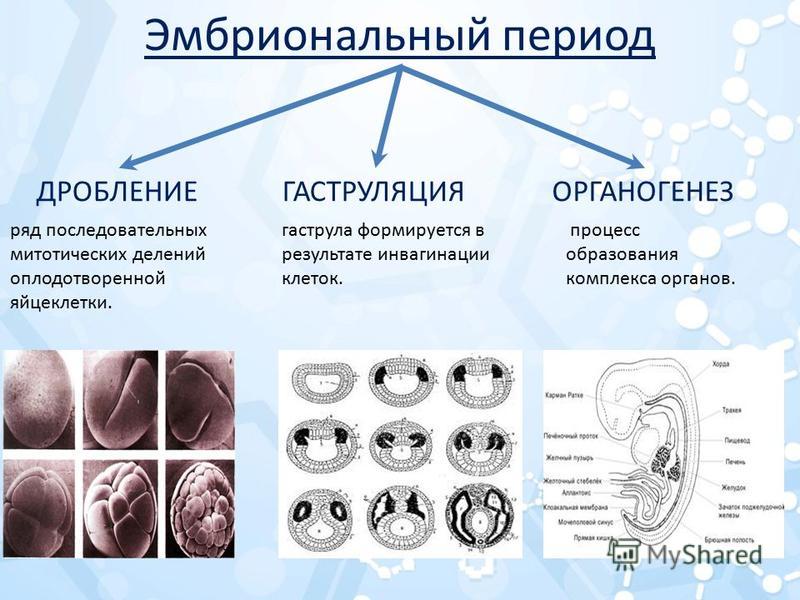
На будущей спинной стороне зародыша начинается впячивание эктодермы в форме желобка — закладывается нервная пластинка, которая в дальнейшем развивается в нервную трубку.
Процесс образования нервной пластинки и её замыкание в нервную трубку в процессе зародышевого развития хордовых называется — нейруляцией.
Нервная трубка погружается под эктодерму и образует зачаток спинного мозга. А из его переднего широкого конца развивается головной мозг.
Таким образом, уже на ранних стадиях эмбрионального периода онтогенеза из внешне одинаковых бластомеров развиваются различные по строению и функциям ткани, органы и системы. Этот процесс получил название дифференцировки клеток.
Дифференцировка
происходит благодаря активации определённых групп генов в различных клетках
зародыша, благодаря чему в клетках синтезируются белки, каждый из которых будет
выполнять те или иные функции.
Следом за эмбриональным развитием сразу наступает постэмбриональное развитие.
У разных организмов этот период может происходить по-разному.
Постэмбриональное развитие начинается с выхода новой особи из яйцевых оболочек (при живорождении) из организма матери.
Постэмбриональное развитие подразделяется на три периода:
ювенИльный период, продолжается до окончания полового созревания;
пубертАтный (период половой зрелости) и период старения.
Развитее организма в ювенильный период.
У разных видов развитие организма в ювенильный период может протекать по двум разным путям.
У одних развитие может быть прямое. У других непрямое.
Прямое
развитие происходит без превращений. В этом случае вновь
появившийся на свет организм отличается от взрослой особи только размерами,
пропорциями и недоразвитием некоторых органов.
Такое развитие наблюдается у рыб, пресмыкающихся, птиц и млекопитающих.
У большинства млекопитающих зародыш развивается внутри тела самки. Такой вариант развития называют внутриутробным.
Когда организм новой особи становится способным к самостоятельному дыханию и питанию, происходят роды — из половых путей самки выходит детёныш.
Непрямое постэмбриональное развитие иногда требует сложной перестройки при переходе к взрослой форме.
Различают два типа непрямого развития — с полным и неполным превращением.
Развитие с превращением характерно для ряда насекомых и земноводных.
Личинки насекомых с неполным превращением внешне сходны со взрослыми особями.
Таким образом, при неполном превращении насекомое проходит три стадии: яйцо, личинка и имаго ― взрослое насекомое.
У
насекомых при развитии с полным превращением особь проходит несколько
последовательных стадий, отличающихся друг от друга образом жизни и характером
питания.
При развитии с превращением из яйца появляется личинка, совершенно не похожая на взрослый организм. Такое развитие называется непрямым или развитием с метаморфозом, т. е. постепенным превращением организма во взрослую особь.
Таким образом, у насекомых с полным развитием в онтогенезе присутствуют четыре стадии: яйцо, личинка, куколка и имаго.
Ювенильный период всегда сопровождается ростом организма. Процесс роста запрограммирован генетически, а также здесь немалую роль играют условия существования.
Пубертатный период — период половой зрелости, у большинства позвоночных животных занимает, большую часть жизни.
После пубертатного периода начинаются изменения, которые снижают возможность организма к приспособлениям к изменяющимся условиям окружающей среды.
Наступает
третий период — период старения. А затем и смерть — прекращение
жизнедеятельности организма. Без смерти не происходила бы смена поколений —
одна из основных движущих сил эволюции.
Без смерти не происходила бы смена поколений —
одна из основных движущих сил эволюции.
Филогенез — это историческое развитие организмов. Филогенез рассматривает эволюцию в качестве процесса, в котором генетическая линия — организмы от предка к потомкам — разветвляется во времени, и её отдельные ветви могут приобретать те или иные изменения или исчезать в результате вымирания. Этот процесс принято изображать графически в виде филогенетического древа, показывающего возможные родственные связи между отдельными ветвями живого.
Этапы онтогенеза | Дистанционные уроки
30-Июл-2013 | комментария 3 | Лолита Окольнова
Онтогенез — индивидуальное развитие организма от оплодотворения (при половом размножении) или от момента отделения от материнской особи (при бесполом размножении) до смерти.

По сути, онтогенез — это путь организма от зачатия и до смерти.
Термин «онтогенез» введен Эрнстом Геккелем в 1866 г.
Индивидуальное развитие (онтогенез) всегда находится под контролем генов.
В процессе онтогенеза реализуется генетическая информация, полученная от родителей.
Все клетки одного организма потенциально несут одну и ту же генетическую программу, но по мере развития организма разные его клетки используют разные части этой программы. На характер работы генов большое влияние оказывают условия внешней среды.
Гаметогенез — развитие половых клеток — гамет до момента, когда они становятся готовыми к оплодотворению.
Партеногенез — развитие организма из неоплодотворенной яйцеклетки.
Эмбриогенез — развитие организма от оплодотворения и до момента выхода из материнского организма (из зародышевой оболочки).
Рост — постэмбриональное развитие — накопление массы, изменение в размерах, становление обмена веществ и т.д. Метаморфозы = превращения. Для некоторых организмов очень важный этап (у бабочек — переход: гусеница — куколка — бабочка)
Старение — процесс постепенного нарушения функций организма, деградация систем организма.
Гаметогенез мы уже рассмотрели.
Перейдем теперь к следующему этапу.
Первый этап онтогенеза — Эмбриогенез
— онтогенез от оплодотворения до рождения
Итак, сперматозойд оплодотворил яйцеклетку. Гаплойдный набор гамет объединился и образовалась диплойдная зигота .
Эта клеточка приступает к интенсивному дроблению — масса и объем зародыша остается постоянным, но внутри — быстрое увеличение (митоз) количества клеток — бластомеров . Образуется бластула.
Образуется бластула.
Так, клеточки поднакопили, пора им форму придать… 🙂
Гаструляция — следующий этап онтогенеза. С делением закончили, теперь клетки интенсивно перемещаются, образуются пласты из этих клеток — зародышевые листки.
Образуется гаструла. Зародыш становится двух- или трехслойным (в зависимости от сложности организма), эти слои в будущем будут формировать органы.
Органогенез. Образование органов и систем органов.
Интересно, что первыми закладываются:
- нервная трубка — будущая ЦНС (центральная нервная система),
- хорда — будущий позвоночник и
- кишечная трубка — будущая пищеварительная система.
Образуется нейрула.
Это самые основные системы любого организма и, как видите, они закладывается уже тогда, когда у зародыша и органов-то еще нет…
Дальше идет накопление массы зародыша, дальнейшая дифференцировка, развитие и т. д.
д.
Постэмбриональный этап онтогенеза
Развитие организма после появления на свет бывает прямое и непрямое.
Прямое развитие — характерно для млекопитающих, рептилий, птиц. Маленький организм отличается от взрослого только размерами и половым созреванием.
Непрямое развитие — развитие с метаморфозами (с превращениями).
Есть такое понятие — имаго — это взрослая особь. На свет появляется личинка, и она может ОЧЕНЬ отличаться от взрослого организма, может иметь такие органы, которые не присущи взрослому. Далее эта личинка претерпевает превращения — метаморфозы (полный или неполный метаморфоз), и уже в результате становится имаго — взрослой особью.
Такое развитие характерно, например, для насекомых и земноводных.
Последующие процессы, такие как рост, будут рассмотрены для каждого типа животных отдельно.
Конечно, у каждого вида организма свое развитие, свой онтогенез. Есть схожие процессы, есть явные различия, но суть остается та же — развитие каждого организма определено генетической информацией, заложенной в половых клетках, которые, объединяясь, дают начало новому организму.
Путь организма от зачатия и до смерти определяется его генетической информацией.
Еще на эту тему:
Обсуждение: «Этапы онтогенеза»
(Правила комментирования)Определение онтогенеза на Dictionary.com
[on-toj-uh-nee] SHOW IPA
/ ɒnˈtɒdʒ ə ni / PHONETIC RESPELLING
существительное Биология.
история развития или развития отдельного организма.
ВИКТОРИНЫ
УЗНАЙТЕ СЕБЯ ИМЕЕТ ПРОТИВ. ИМЕЮТ!
У вас есть грамматические навыки, чтобы знать, когда использовать «иметь» или «иметь»? Давайте узнаем это с помощью этой викторины!
Вопрос 1 из 7
Моя бабушка ________ Стена, полная старинных часов с кукушкой.
Происхождение онтогенеза
Впервые зарегистрировано в 1870–75; on- + -genyДРУГИЕ СЛОВА ИЗ онтогенеза
on · to · ge · net · ic [on-tuh-juh-net-ik], / ˌɒn tə dʒəˈnɛt ɪk /, on · to · ge · net · i · cal , на · to · gen · ic, прилагательное · to · ge · net · i · cal·ly, on · to · gen · i · cal·ly, наречие · tog · e · nist, существительноеСлова рядом с онтогенезом
на на цыпочках, на, на функцию, онтогенез, онтогенетика, онтогенез, онтологический, онтологический аргумент, онтологизм, онтологизация, онтологияСловарь.com Несокращенный На основе Несокращенного словаря Random House, © Random House, Inc. 2021
Примеры предложений из Интернета для онтогенеза
-
Это выражено формулой Герберта Спенсера: «Онтогенез повторяет филогенез».
-
Он опирается на факты палеонтологии и заполняет пробелы сравнительной анатомией и онтогенезом.
-
Онтогенез безошибочно учит, что его элементарные органы, клетки эпидермиса, полностью развиваются из эктодермы.

-
Три наиболее ценных источника данных по филогении — это палеонтология, сравнительная анатомия и онтогенез.
онтогенез
онтогенез (ˌɒntəˈdʒɛnɪsɪs)
существительное
полная последовательность событий, участвующих в развитии 9002 флогенических форм отдельных организмов онтогенетический (ˌɒntəˈdʒɛnɪk) или онтогенетический (ˌɒntədʒɪˈnɛtɪk), прилагательное онтогенетически или онтогенетически, наречиеCollins English Dictionary — Complete & Unabridged 2012 Digital Edition © William Collins Sons & Co.Ltd. 1979, 1986 © HarperCollins Издательство 1998, 2000, 2003, 2005, 2006, 2007, 2009, 2012
н.
Происхождение и развитие отдельного организма от эмбриона до взрослого. Ontogenesis
Медицинский словарь Стедмана American Heritage®
Авторское право © 2002, 2001, 1995 компанией Houghton Mifflin. Опубликовано компанией Houghton Mifflin.
Опубликовано компанией Houghton Mifflin.
Происхождение и развитие отдельного организма от эмбриона до взрослого.
Научный словарь американского наследия® Авторские права © 2011.Опубликовано Houghton Mifflin Harcourt Publishing Company. Все права защищены.
Онтогенез | Encyclopedia.com
Онтогенез — это теория развития и структурирования индивида, которая принимает во внимание происхождение индивида и условия его или ее развития. Зигмунд Фрейд всегда связывал онтогенез человека с филогенезом, принимая биогенетическую точку зрения.
Онтогенез включает в себя процессы развития и приобретения, специфичные для индивидуума, в отличие от филогенеза, который включает процессы эволюции и приобретения, специфичные для вида.Онтогенез и филогенез не независимы друг от друга: индивидуальные приобретения возможны только в установленных пределах вида.
Определение онтогенеза, или развития индивида, требует изучения того, что составляет разум и каковы основные условия его организации. Требуется не что иное, как четкое понимание происхождения, стадий развития и более ранних состояний истории человека.
Требуется не что иное, как четкое понимание происхождения, стадий развития и более ранних состояний истории человека.
В конце девятнадцатого века психология нашла плодородную почву в всеобъемлющей теории эволюции видов животных Дарвина.В то время считалось, что уровень функционирования более ранних видов сохранялся в виртуальном состоянии у более поздних видов. Как сказал Эрнст Геккель: «Онтогенез повторяет филогению». Геккель и другие девятнадцатого века Мыслители-эволюционисты считали, что у людей развитие плода до взрослой жизни (онтогенез), вкратце, соответствует всей истории вида (филогенез).
Между тем британский невролог Джон Хьюлингс Джексон выдвинул гипотезу об иерархической организации центральной нервной системы, показав, что приостановка контроля со стороны высших центров высвобождает архаические автоматические реакции в низших центрах, которые соответствуют более ранним фазам развития.Фрейд черпал вдохновение из этой модели с понятием «регрессия». Во Франции Анри Эй основал свою теорию органодинамизма на «растворениях» Джексона.
В то время преобладала идея, что некоторые функции, имеющие адаптивную ценность для низших видов животных, сохраняются в виртуальном состоянии и могут быть реактуализированы патологией: отсутствие контроля на более высоком уровне высвобождает более низкие уровни иерархии. Идея о том, что патология высвобождает менее дифференцированный, более «автоматический» уровень функционирования, восходящий к более ранней стадии индивидуального или видового развития, оказала влияние на многих авторов, включая Фрейда и Пьера Жане.
Таким образом, Фрейд в своих самых ранних работах неявно принял биогенетическую концепцию полового развития человека. Вместе с Вильгельмом Стекелем, Вильгельмом Бёльше, Гранвиллом Стэнли Холлом и многими другими он применил «фундаментальный биогенетический закон» Геккеля к вопросу о половом развитии.
Если ребенок в процессе своего роста повторяет историю вида, Фрейд пришел к выводу, что он должен, в более широком смысле, повторять половую историю вида.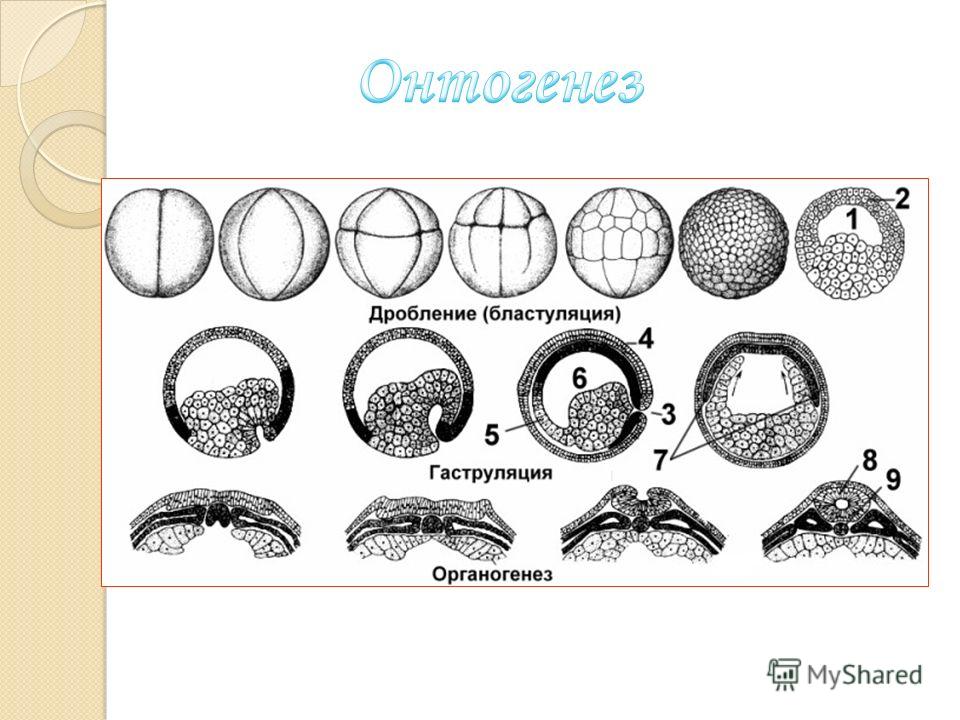 Другими словами, препубертатный человек должен обладать врожденной способностью испытывать все архаичные формы сексуального удовольствия, которые характеризовали взрослые стадии наших далеких предков.В «Некоторые мысли о развитии и регрессе — этиология», двадцать вторая из его вводных лекций (1916-1917a), Фрейд писал о развитии эго и либидо, что «оба они являются в основе наследства, сокращенными повторениями. о развитии, которое все человечество прошло с первых дней своего существования в течение длительного периода времени »(стр. 354).
Другими словами, препубертатный человек должен обладать врожденной способностью испытывать все архаичные формы сексуального удовольствия, которые характеризовали взрослые стадии наших далеких предков.В «Некоторые мысли о развитии и регрессе — этиология», двадцать вторая из его вводных лекций (1916-1917a), Фрейд писал о развитии эго и либидо, что «оба они являются в основе наследства, сокращенными повторениями. о развитии, которое все человечество прошло с первых дней своего существования в течение длительного периода времени »(стр. 354).
Эта биогенетическая логика не только подтвердила мысли Фрейда о «полиморфно перверсивной» природе детской сексуальности, но также позже подкрепила другие разработки его теории.Понятия «фиксация» (задержка или задержка развития), «регресс» (связана с общим понятием «растворение» Джексона) и «персеверация ранних впечатлений» (после модели эмбриологических экспериментов Вильгельма Ру) — все они возникли с точки зрения развития. чтобы объяснить организацию неврозов.
Фрейд следовал логике повторения в сноске, добавленной в 1915 году к его Три эссе по теории сексуальности (1905d), где он настаивал на том, что каждая основная «прегенитальная» стадия детского полового развития сохраняет специфическое унаследование от этого филогенетического влияние.С той же точки зрения, психическая патология считалась возвращением прошлых стадий онтогенетического и филогенетического развития, которые интегрированы и в то же время открыты заново. Для Фрейда формирование симптомов актуализировало прошлое, а психоаналитическое лечение само по себе было терапевтической формой воспоминаний.
В поисках истоков эдипова конфликта Фрейд подробно описал серию из трех первичных фантазий — соблазнения, кастрации и первичной сцены, — которые он считал данностью с самого начала и лежащими в основе всей психической организации, помимо травмирующих событий. истории человека.Предполагаемая универсальность Эдипова комплекса привела его к мысли, что в каждом индивидууме существует эмбриональная версия этих фантазий, которые встречаются во всех культурах, как филогенетически переданный след убийства отца первобытной ордой.
Неоламаркистская перспектива Фрейда и его филогенетические теории вызвали немалую критику, несмотря на исследование Фрэнка Дж. Саллоуэя « Фрейд — биолог разума » (1979), в котором был представлен весь объем построений Фрейда.Дело в том, что в начале двадцать первого века культурная и языковая передача, наряду с коммуникацией тем или иным способом, из родительского или дедовского или дедовского бессознательного, гораздо охотнее используется для определения способа происхождения самых ранних конститутивных фантазий психики, чем гипотетическая запись в генетическом материале вида. Исследование трансгенерационных эффектов (Kaës, 1993) идет в этом направлении, как и «общая теория соблазнения» Жана Лапланша, которая утверждает, что материнское бессознательное передает сексуализированные сообщения или «загадочные означающие» ребенку.
Современные исследования связаны не столько с открытием окончательных истоков формирования личности, сколько с объяснением условий, в которых это происходит. С 2005 г. онтогенетический подход, то есть исследование развития индивида или генезиса
Эдипов комплекс — концентрируется на двух основных направлениях исследования. Первый — либидинозное развитие, принимая во внимание последовательные фазы психической организации, с акцентом на преобладающую эрогенную зону, вокруг которой сосредоточено психическое возбуждение и организуются фантазии.Вторая направлена на развитие объектных отношений в их взаимосвязи с развитием эго.
С 2005 г. онтогенетический подход, то есть исследование развития индивида или генезиса
Эдипов комплекс — концентрируется на двух основных направлениях исследования. Первый — либидинозное развитие, принимая во внимание последовательные фазы психической организации, с акцентом на преобладающую эрогенную зону, вокруг которой сосредоточено психическое возбуждение и организуются фантазии.Вторая направлена на развитие объектных отношений в их взаимосвязи с развитием эго.
Жан-Франсуа Рабен
См. Также: Цивилизация и ее недовольство ; Претензии психоанализа на научный интерес; культурная передача; Дарвин, дарвинизм и психоанализ; Отцовство; Процессы развития; Наследственность приобретенных характеров; Межпоколенческие; Недостаток дифференциации; Материнская мечтательность, способность к; Филогенез; Предыстория; Первобытная орда; Thalassa.Теория генитальности ; Тотем и табу .
Библиография
Фрейд, Зигмунд, (1905d) Три очерка по теории сексуальности. SE , 7: 123-243.
SE , 7: 123-243.
——. (1916-1917а). Вводные лекции по психоанализу. SE , 16.
Каэс, Рене (1993). Вступление. В Рене Каэс и др., Transmission de la vie mental entre générations . Пэрис: Данод.
Саллоуэй, Фрэнк Дж. (1979). Фрейд — биолог разума. Лондон: Бернетт.
Значение онтогенеза
| Участок эукариотической клетки с ее выступающим ядром, в котором запутаны длинные молекулы ДНК (носители генетических инструкций). Другие важные клеточные компоненты или органеллы плавают в эндоплазматической жидкости между ядром и внешней стенкой клетки. Эти органеллы включают митохондрии (отвечающие за энергетический баланс), лизосомы (пищеварение), ретикулум и аппарат Гольджи.Человеческое тело состоит из эукариотических клеток, их около 100000000000. Сравнение эмбрионов позвоночных на одной и той же стадии развития. (Предоставлено: Г. Романес) |
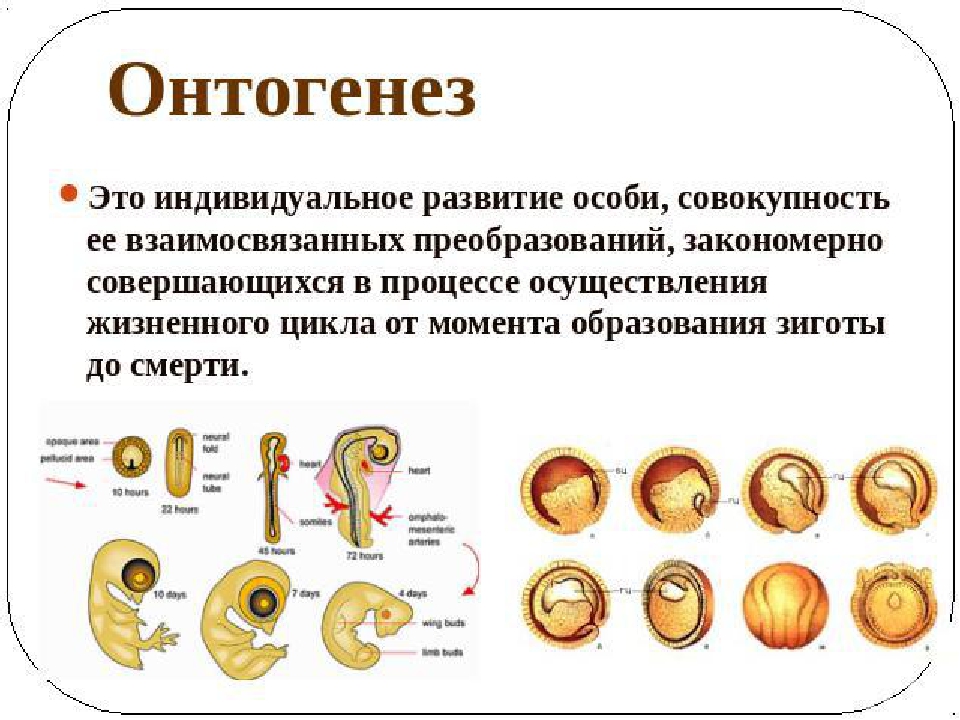 Первые 100 миллионов лет или около того были отмечены сильным таянием и переплавлением горных пород на поверхности планеты. После того, как этот процесс замедлился, начала образовываться твердая корка; но непрерывная бомбардировка комет и астероидов продолжала разрывать его лицо еще несколько сотен миллионов лет.Изменения на поверхности, которые начали происходить в это время, привели к созданию среды, способной поддерживать органические соединения. Вода выделялась из горячих пород под коркой (процесс, называемый «дегазация»). Этот процесс продолжался какое-то время и может все еще быть в какой-то степени активным. Однако вода, выбрасываемая сегодня из вулканов, гейзеров и подводных жерл, в основном представляет собой переработанную поверхностную воду.
Первые 100 миллионов лет или около того были отмечены сильным таянием и переплавлением горных пород на поверхности планеты. После того, как этот процесс замедлился, начала образовываться твердая корка; но непрерывная бомбардировка комет и астероидов продолжала разрывать его лицо еще несколько сотен миллионов лет.Изменения на поверхности, которые начали происходить в это время, привели к созданию среды, способной поддерживать органические соединения. Вода выделялась из горячих пород под коркой (процесс, называемый «дегазация»). Этот процесс продолжался какое-то время и может все еще быть в какой-то степени активным. Однако вода, выбрасываемая сегодня из вулканов, гейзеров и подводных жерл, в основном представляет собой переработанную поверхностную воду. «Известно, что крылья птиц и летучих мышей, а также ноги лошадей и других четвероногих неразличимы в ранний эмбриональный период и что они становятся различимыми по незаметно тонким шагам.Так пишет Дарвин (6-е изд., Стр. 226), используя это наблюдение относительно индивидуального развития организмов в защиту постепенной эволюции по наследственным линиям.
Индивидуальное развитие организма называется онтогенезом. Его линия предков определяет его филогенез. Есть ли между ними связь? Действительно ли онтогенез дает ключ к разгадке происхождения, как утверждает Дарвин?
Самая успешная группа организмов на этой планете, когда в качестве критерия используется видовое разнообразие, — это насекомые.Они вторглись в каждый укромный уголок земного царства, включая пресноводные водоемы. Мы можем подумать, что жуков много, но их личинок «личинок» еще больше, за исключением того, что они спрятаны в земле. У многих других многоклеточных организмов есть незрелые стадии, которые сильно отличаются от зрелых. Помимо личинок насекомых, головастики лягушек, возможно, являются наиболее известными примерами для нас, жителей суши.
В океане личинки прибрежных организмов населяют прибрежный океан в виде микроскопического «меропланктона».Меропланктон состоит из личинок кораллов, морских звезд, многощетинковых червей, ракушек и крабов, мидий и улиток и многих других. Именно по таким личинкам (совсем не похожим на взрослые стадии) можно легко сказать, что ракушки и крабы принадлежат к одному и тому же генеральному племени животных. То же самое и с мидиями и улитками.
Именно по таким личинкам (совсем не похожим на взрослые стадии) можно легко сказать, что ракушки и крабы принадлежат к одному и тому же генеральному племени животных. То же самое и с мидиями и улитками.
С точки зрения функции, большие различия между личиночной стадией и половозрелым животным очевидны. Здесь задействованы две стратегии, отвечающие двум различным потребностям.Один — это поиск ресурсов, а другой — рассеивание. У насекомых с четко выраженным личиночным развитием личинки отвечают за питание и прибавку в весе, в то время как основная задача крылатых стадий — поиск партнеров и наблюдение за расселением. (Дополнительный корм используется для поддержания и производства большего количества яиц). В прибрежных водах океана сидячие взрослые (постоянно прикрепленные) стадии отвечают за рост и размножение, в то время как основная функция личинок — это расселение, то есть поиск полезных ископаемых. места для поселения.(Конечно, личинки также питаются, чтобы поддерживать свой метаболизм и получить хорошую фору при заселении. ) У лягушек и личинки, и взрослые животные питаются и растут, но они используют разные среды обитания: одна в воде, другая — в воде. его краю или за его пределами.
) У лягушек и личинки, и взрослые животные питаются и растут, но они используют разные среды обитания: одна в воде, другая — в воде. его краю или за его пределами.
Тот факт, что личинки многих отдаленно родственных организмов больше похожи друг на друга, чем взрослые животные (например, ракушки и крабы; оболочники, головастики и рыбы), в дополнение к наблюдениям, касающимся эмбрионов позвоночных (цитированных выше), привлек внимание к возможность того, что в развитии есть ключи к родству и, следовательно, к эволюции.Эта принципиально правильная идея породила броскую фразу «онтогенез повторяет филогенез», подразумевающую, что развитие повторяет аспекты этапов эволюции в пределах одного организма. Буквально это чушь; ни человеческие эмбрионы, ни эмбрионы домашней птицы не имеют стадии рыбы. Однако, когда его воспринимают как приглашение искать ключи к разгадке хода эволюции, это имеет смысл. Эмбрионы человека, куриные эмбрионы и эмбрионы рыб имеют много общего на ранних стадиях своего развития, как показано на рисунке выше.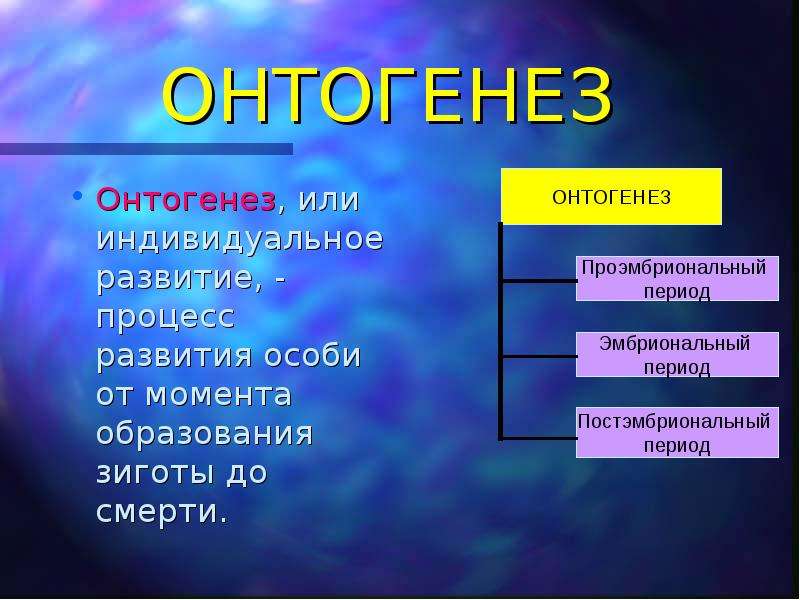
Немецкий биолог Эрнст Геккель (1834-1919) больше, чем кто-либо другой, настаивал на важности онтогенеза как ключа к «Древу жизни». Это его самый известный вклад в науку о жизни. (В свое время он также был мировым экспертом по радиоляриям, известковым губкам и кишечнополостным. Властный писатель и лектор, он убеждал своих коллег и общественность в теории эволюции.) По его мнению, причина того, что онтогенез протекает именно так делает то, что каждый организм помнит свою эволюцию.Раннее развитие обусловлено древними программами; чем раньше разработка, тем старше программа.
Верно ли это для какого-либо конкретного организма, нельзя решить путем спора. Ответ придет из изучения генетического кода (концепция, недоступная во времена Геккеля). Поскольку эволюционное давление действует как на личинок, так и на взрослых особей (личинки должны искать пищу и спасаться от хищников), мы должны ожидать обнаружения изменений в генетическом коде для улучшения адаптации личинок.У эмбрионов есть и другие проблемы: они должны убедить родителей, что им нужно больше ресурсов. Соответствующие сигналы предположительно будут разными для акул, птиц и млекопитающих, что приведет к расхождению программ развития. Из-за этих оговорок концепция «онтогенез повторяет филогенез» не может считаться общим правилом.
Соответствующие сигналы предположительно будут разными для акул, птиц и млекопитающих, что приведет к расхождению программ развития. Из-за этих оговорок концепция «онтогенез повторяет филогенез» не может считаться общим правилом.
В любом случае закономерности развития животных, растений и грибов широко использовались для создания классификаций, которые, как считается, отражают родственные связи с точки зрения общего происхождения.Ясно, что всякий раз, когда два разных организма имеют сложные, но похожие программы развития, следует предполагать, что они унаследовали свои программы от общего предка, а не то, что программы возникли независимо, случайно.
Онтогенез и филогения: молекулярные признаки отбора, ограничения и временной плейотропии в развитии Drosophila | BMC Biology
Von Baer KE: Entwicklungsgeschichte der Tiere: Beobachtung und Relexion. 1828, Кенигсберг, Германия: Bornträger
Google ученый
Рейдл Р: Порядок в живых организмах. 1978, Нью-Йорк, США: John Wiley & Sons
Google ученый
Ричардсон М.К., Ханкен Дж., Селвуд Л., Райт Дж. М., Ричардс Р. Дж., Пио С., Рейно А. Геккель, эмбрионы и эволюция. Наука. 1998, 280: 983-986.10.1126 / science.280.5366.983c.
Артикул CAS PubMed Google ученый
Рафф Р.А.: Форма жизни: развитие генов и эволюция формы животных. 1996, Чикаго, Иллинойс, США: The University of Chicago Press
Google ученый
Сандер К. Эволюция механизмов формирования паттерна: извлечение из эмбриогенеза и сперматогенеза насекомых. Развитие и эволюция. Отредактировано: Goodwin BC, Holder N, Wylie CC. 1983, Кембридж, Массачусетс, США: Cambridge University Press, 137–159.
Google ученый
Галис Ф., Мец Дж. А.: Проверка уязвимости филотипической стадии: модульность и эволюционное сохранение.J Exp Zoo. 2001, 291: 195-204. 10.1002 / jez.1069.
Артикул CAS Google ученый
Дарвин К. Происхождение человека и отбор в отношении пола.1871 г., Принстон, штат Нью-Джерси. США: Princeton University Press
Google ученый
Дарвин С: О происхождении видов посредством естественного отбора или о сохранении привилегированных рас в борьбе за жизнь. 1872, Нью-Йорк, США: Современная библиотека, 6
Google ученый
Артур В .: Теория эволюции развития. 1988, Нью-Йорк, США: John Wiley & Sons
Google ученый
Каттер А.Д., Уорд С.: Сексуальная и временная динамика молекулярной эволюции в развитии C. elegans . Mol Biol Evol. 2005, 22: 178-188. 10.1093 / молбев / мш367.
Артикул CAS PubMed Google ученый
Дэвидсон Э. Х., Макклей Д. Р., Худ Л.: Регуляторные генные сети и свойства процесса развития. Proc Natl Acad Sci USA. 2003, 100: 1475-1480. 10.1073 / pnas.0437746100.
Proc Natl Acad Sci USA. 2003, 100: 1475-1480. 10.1073 / pnas.0437746100.
PubMed Central Статья CAS PubMed Google ученый
Wittkopp PJ: Экспрессия вариабельных генов у эукариот: сетевая перспектива. J Exp Biol. 2007, 210: 1567-1575. 10.1242 / jeb.002592.
Артикул CAS PubMed Google ученый
Дэвидсон Э.Х., Эрвин Д.Х.: Сети регуляции генов и эволюция планов тела животных.Наука. 2006, 311: 796-800. 10.1126 / science.1113832.
Артикул CAS PubMed Google ученый
Lewis EB: генный комплекс, контролирующий сегментацию у Drosophila . Природа. 1978, 276: 565-570. 10.1038 / 276565a0.
Природа. 1978, 276: 565-570. 10.1038 / 276565a0.
Артикул CAS PubMed Google ученый
Lutz B, Lu HC, Eichele G, Miller D, Kaufman TC: Спасение лабиального нуль-мутанта Drosophila с помощью куриного ортолога Hoxb-1 демонстрирует, что функция Hox-генов сохраняется филогенетически.Genes Dev. 1996, 10: 176-184. 10.1101 / gad.10.2.176.
Артикул CAS PubMed Google ученый
Лимонов Д., МакГиннис В. Геномная эволюция кластеров генов Hox. Наука. 2006, 313: 1918-1922. 10.1126 / science.1132040.
Артикул CAS PubMed Google ученый
Хинман В.Ф., Нгуен А.Т., Камерон Р.А., Дэвидсон Э.Х .: Архитектура регуляторной сети генов развития на протяжении 500 миллионов лет эволюции иглокожих. Proc Natl Acad Sci USA. 2003, 100: 13356-13361. 10.1073 / pnas.2235868100.
PubMed Central Статья CAS PubMed Google ученый
Кастильо-Дэвис К.И., Хартл Д.Л.: Эволюция генома и ограничения развития у Caenorhabditis elegans . Mol Biol Evol. 2002, 19: 728-735.
Артикул CAS PubMed Google ученый
Дэвис Дж. С., Брандман О., Петров Д. А.: Эволюция белков в контексте развития Drosophila . J Mol Evol. 2005, 60: 774-785. 10.1007 / s00239-004-0241-2.
Артикул CAS PubMed Google ученый
Hill AA, Hunter CP, Tsung BT, Tucker-Kellogg G, Brown EL: Геномный анализ экспрессии генов в C. elegans . Наука. 2000, 290: 809-812. 10.1126 / science.290.5492.809.
Артикул CAS PubMed Google ученый
Reinke V, Gil IS, Ward S, Kazmer N: обогащенные по всей зародышевой линии и смещенные по полу профили экспрессии в C. elegans . Mol Cell. 2004, 6: 605-616. 10.1016 / S1097-2765 (00) 00059-9.
Артикул Google ученый
Арбайтман М.Н., Ферлонг Е.Е., Имам Ф., Джонсон Е., Нулевой Б.Х., Бейкер Б.С., Краснов М.А., Скотт М.П., Дэвис Р.В., Уайт КП: экспрессия генов в течение жизненного цикла Drosophila melanogaster . Наука. 2002, 297: 2270-2275. 10.1126 / science.1072152.
Наука. 2002, 297: 2270-2275. 10.1126 / science.1072152.
Артикул CAS PubMed Google ученый
Musters H, Huntley MA, Singh RS: геномное сравнение эволюции более быстрого пола, быстрого X и более быстрого самца между Drosophila melanogaster и Drosophila pseudoobscura .J Mol Evol. 2006, 62: 693-700. 10.1007 / s00239-005-0165-5.
Артикул CAS PubMed Google ученый
Graur D, Li WH: Основы молекулярной эволюции. 2002, Сандерленд, Коннектикут, США: Sinauer
Google ученый
Zhang Y, Sturgill D, Parisi M, Kumar S, Oliver B: Ограничение и оборот в экспрессии генов, зависимых от пола, в роде Drosophila . Природа. 2007, 450: 233-237. 10.1038 / природа06323.
Природа. 2007, 450: 233-237. 10.1038 / природа06323.
PubMed Central Статья CAS PubMed Google ученый
Drosophila 12 Genomes Consortium: Эволюция генов и геномов в филогении Drosophila . Природа. 2007, 450: 203-218. 10.1038 / природа06341.
Артикул Google ученый
Lachaise D, Cariou ML, David JR, Lemeunier F, Tsacas L, Ashburner M: Историческая биогеография подгруппы видов Drosophila melanogaster .Evol Biol. 1988, 22: 159-226.
Google ученый
Понтиус Ю.Ю., Вагнер Л., Шулер Г.Д .: UniGene: единый взгляд на транскриптом. Справочник NCBI. 2003, Бетесда, Мэриленд, США: Национальный центр биотехнологической информации
Справочник NCBI. 2003, Бетесда, Мэриленд, США: Национальный центр биотехнологической информации
Google ученый
Al-Shahrour F, Minguez P, Tárraga J, Medina I, Alloza E, Montaner D, Dopazo J: FatiGO +: инструмент функционального профилирования геномных данных.Интеграция функциональной аннотации, регуляторных мотивов и данных взаимодействия с экспериментами на микрочипах. Nucleic Acids Res. 2007, 35: W91-6. 10.1093 / нар / гкм260.
PubMed Central Статья PubMed Google ученый
Аль-Шахрур Ф., Диас-Уриарте Р., Допазо Дж .: FatiGO: веб-инструмент для поиска значимых ассоциаций терминов генной онтологии с группами генов. Биоинформатика. 2004, 20: 578-580. 10.1093 / биоинформатика / btg455.
Артикул CAS PubMed Google ученый
Vicario S, Mason CE, White KP, Powell JR: Стадия развития и уровень систематической ошибки использования кодонов у Drosophila . Mol Biol Evol. 2008, 25: 2269-2277. 10.1093 / molbev / msn189.
PubMed Central Статья CAS PubMed Google ученый
Хартенштейн V: Атлас развития дрозофилы.1993, Колд-Спринг-Харбор, Нью-Йорк, США: Лабораторный пресс Колд-Спринг-Харбор
Google ученый
Хайтович П., Хеллманн И., Энард В., Новик К., Лайнвебер М., Франц Х., Вайс Г., Лахманн М., Пяабо С. Параллельные модели эволюции геномов и транскриптомов человека и шимпанзе. Наука. 2005, 309: 1850-1854. 10.1126 / science.1108296.
Параллельные модели эволюции геномов и транскриптомов человека и шимпанзе. Наука. 2005, 309: 1850-1854. 10.1126 / science.1108296.
Артикул CAS PubMed Google ученый
Haerty W, Jagadeeshan S, Kulathinal RJ, Wong A, Ravi Ram K, Sirot LK, Levesque L, Artieri CG, Wolfner MF, Civetta A и др.: Эволюция в быстром темпе: быстро эволюционирующие гены, связанные с полом, у Drosophila . Генетика. 2007, 177: 1321-1335. 10.1534 / genetics.107.078865.
PubMed Central Статья CAS PubMed Google ученый
Ларракуэнте А.М., Сактон ТБ, Гринберг А.Дж., Вонг А., Сингх Н.Д., Стерджилл Д., Чжан И., Оливер Б., Кларк А.Г.: Эволюция генов, кодирующих белок, в Drosophila . Тенденции Genet. 2008, 24: 114-123. 10.1016 / j.tig.2007.12.001.
Тенденции Genet. 2008, 24: 114-123. 10.1016 / j.tig.2007.12.001.
Артикул CAS PubMed Google ученый
Good JM, Nachman MW: Скорость эволюции белка положительно коррелирует со временем развития экспрессии во время сперматогенеза мыши. Mol Biol Evol. 2005, 22: 1044-1052. 10.1093 / molbev / msi087.
Артикул CAS PubMed Google ученый
Янг З., Нильсен Р. Оценка синонимичных и несинонимичных коэффициентов замещения в реалистичных эволюционных моделях. Mol Biol Evol. 2000, 17: 32-43.
Артикул CAS PubMed Google ученый

Shigenobu S, Kitadate Y, Noda C, Kobayashi S: Молекулярная характеристика эмбриональных гонад с помощью профилирования экспрессии генов в Drosophila melanogaster . Proc Natl Acad Sci USA. 2006, 12: 13728-13733. 10.1073 / pnas.0603767103.
Артикул Google ученый
Райт TRF, (ред.): Генетические регуляторные иерархии в развитии. 1990, Торонто, Канада: Academic Press
Google ученый
Старк С., Брайткройц Б.Дж., Регули Т., Бушер Л., Брейткройц А., Тайерс М.: BioGRID: общий репозиторий для наборов данных взаимодействия. Nucleic Acids Res. 2006, 34: D535-9. 10.1093 / нар / gkj109.
PubMed Central Статья CAS PubMed Google ученый
Fraser HB: Модульность и эволюционные ограничения белков.Нат Жене. 2005, 37: 351-352. 10.1038 / ng1530.
Артикул CAS PubMed Google ученый
Lemos B, Bettencourt BR, Meiklejohn CD, Hartl DL: Эволюция белков и уровни экспрессии генов связаны в Drosophila и независимо связаны с количеством мРНК, длиной белка и количеством белок-белковых взаимодействий. Mol Biol Evol. 2005, 22: 1345-1354.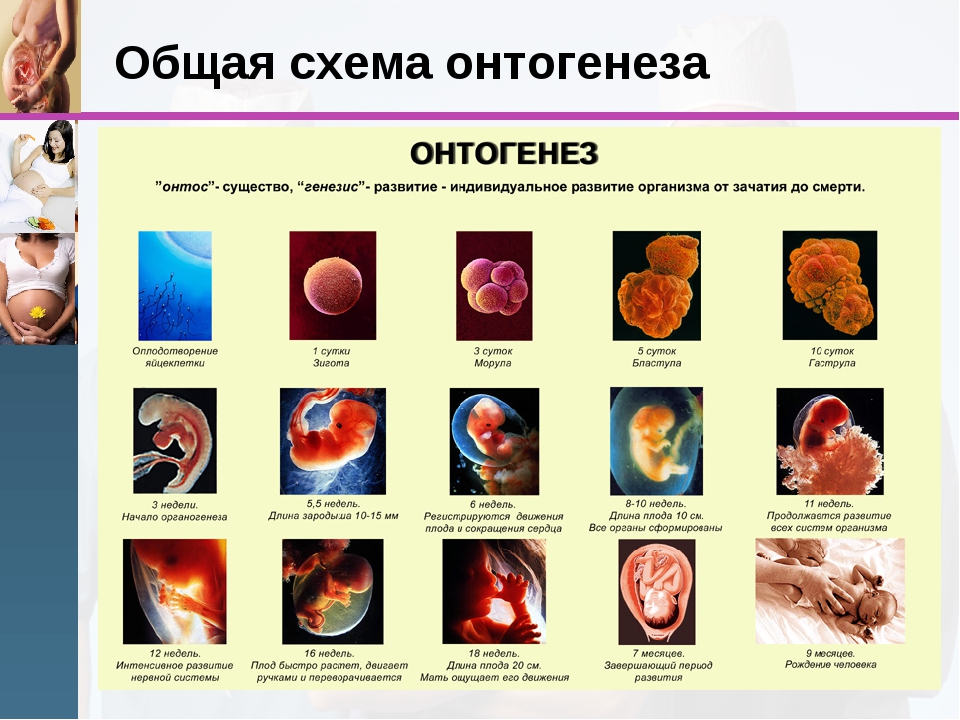 10.1093 / molbev / msi122.
10.1093 / molbev / msi122.
Артикул CAS PubMed Google ученый
Ranz JM, Castillo-Davis CI, Meiklejohn CD, Hartl DL: Зависимая от пола экспрессия генов и эволюция транскриптома Drosophila . Наука. 2003, 300: 1742-1745. 10.1126 / science.1085881.
Артикул CAS PubMed Google ученый
Meiklejohn CD, Parsch J, Ranz JM, Hartl DL: Быстрая эволюция экспрессии генов с предвзятостью самцов в Drosophila . Proc Natl Acad Sci USA. 2003, 100: 9894-9899.10.1073 / pnas.16306.
PubMed Central Статья CAS PubMed Google ученый
Акаши H: Экспрессия генов и молекулярная эволюция.Curr Opin Genet Dev. 2001, 11: 660-666. 10.1016 / S0959-437X (00) 00250-1.
Артикул CAS PubMed Google ученый
Sharp PM, Li WH: Индекс адаптации кодонов — мера направленного смещения использования синонимичных кодонов и его потенциальных приложений. Nucleic Acids Res. 1987, 15: 1281-1295. 10.1093 / нар / 15.3.1281.
PubMed Central Статья CAS PubMed Google ученый
Stanyon CA, Liu G, Mangiola BA, Patel N, Giot L, Kuang B, Zhang H, Zhong J, Finley RL: A Drosophila protein- карта взаимодействия сосредоточена на регуляторах клеточного цикла. Genome Biol. 2004, 5: R96-10.1186 / gb-2004-5-12-r96.
PubMed Central Статья PubMed Google ученый
Харт Г.Т., Рамани А.К., Маркотт Э.М.: Насколько полны современные дрожжевые и человеческие сети взаимодействия белков ?.Genome Biol. 2006, 7: 120-10.1186 / GB-2006-7-11-120.
PubMed Central Статья PubMed Google ученый

Чиветта А., Сингх RS: Пол и видообразование: генетическая архитектура и эволюционный потенциал половых и несексуальных признаков у видов-братьев из комплекса Drosophila melanogaster . Эволюция. 1998, 52: 1080-1092. 10.2307 / 2411238.
Артикул Google ученый
Civetta A, Singh RS: Высокая дивергенция белков репродуктивного тракта и их связь с постзиготической репродуктивной изоляцией у видов группы Drosophila melanogaster и Drosophila virilis . J Mol Evol. 1995, 41: 1085-1095. 10.1007 / BF00173190.
Артикул CAS PubMed Google ученый
Swanson WJ, Vacquier VD: Быстрая эволюция репродуктивных белков. Nat Rev Genet. 2002, 3: 137-144.10.1038 / nrg733.
Артикул CAS PubMed Google ученый
Сингх Р.С., Кулатинал Р.Дж.: Эволюция и видообразование полового генофонда — новая парадигма. Genes Genetic Sys. 2000, 75: 119-130. 10.1266 / ggs.75.119.
Артикул CAS Google ученый
Сингх Р.С., Кулатинал Р.Дж.: Мужское половое влечение и маскулинизация генома. Биологические исследования. 2005, 27: 518-525.10.1002 / bies.20212.
Артикул CAS PubMed Google ученый
Артьери К.Г., Хэрти В., Гупта Б.П., Сингх Р.С.: Половой отбор и поддержание пола: данные сравнения темпов геномного накопления мутаций и дивергенции связанных с полом генов у половых и гермафродитных видов Caenorhabditis . Mol Biol Evol. 2008, 25: 972-979. 10.1093 / molbev / msn046.
Mol Biol Evol. 2008, 25: 972-979. 10.1093 / molbev / msn046.
Артикул CAS PubMed Google ученый
Pröschel M, Zhang Z, Parsch J: Широко распространенная адаптивная эволюция генов Drosophila со смещенной по полу экспрессией. Генетика. 2006, 174: 893-900. 10.1534 / genetics.106.058008.
PubMed Central Статья PubMed Google ученый
Бейнс Дж. Ф., Сойер С. А., Хартл Д. Л., Парш Дж.: Влияние х-сцепления и смещенной по полу экспрессии генов на скорость эволюции адаптивного белка у Drosophila . Mol Biol Evol.2008, 25: 1639-1650. 10.1093 / молбев / msn111.
PubMed Central Статья CAS PubMed Google ученый
Cruickshank T, Wade MJ: Микроэволюционная поддержка «песочных часов» развития: вариация и дивергенция формы паттернов экспрессии генов в Drosophila .Evol Dev. 2008, 10: 583-590. 10.1111 / j.1525-142X.2008.00273.x.
Артикул PubMed Google ученый
He X, Zhang J: К молекулярному пониманию плейотропии. Генетика. 2006, 173: 1885–1891. 10.1534 / genetics.106.060269.
PubMed Central Статья CAS PubMed Google ученый
Flybase: база данных генов и геномов дрозофилы . [http://flybase.org/]
Babelomics v3.1. [http://babelomics.bioinfo.cipf.es/]
The BioGRID: общий репозиторий для наборов данных взаимодействия. [http://www.thebiogrid.org]
R Development Core Team: R: язык и среда для статистических вычислений. 2004, Вена, Австрия: Фонд R для статистических вычислений
Google ученый